

Activation of EPAC1/2 is essential for osteoclast formation by modulating NFκB nuclear translocation and actin cytoskeleton rearrangements
- PMID: 25122553
- PMCID: PMC4200330
- DOI: 10.1096/fj.14-255703
Activation of EPAC1/2 is essential for osteoclast formation by modulating NFκB nuclear translocation and actin cytoskeleton rearrangements
Abstract
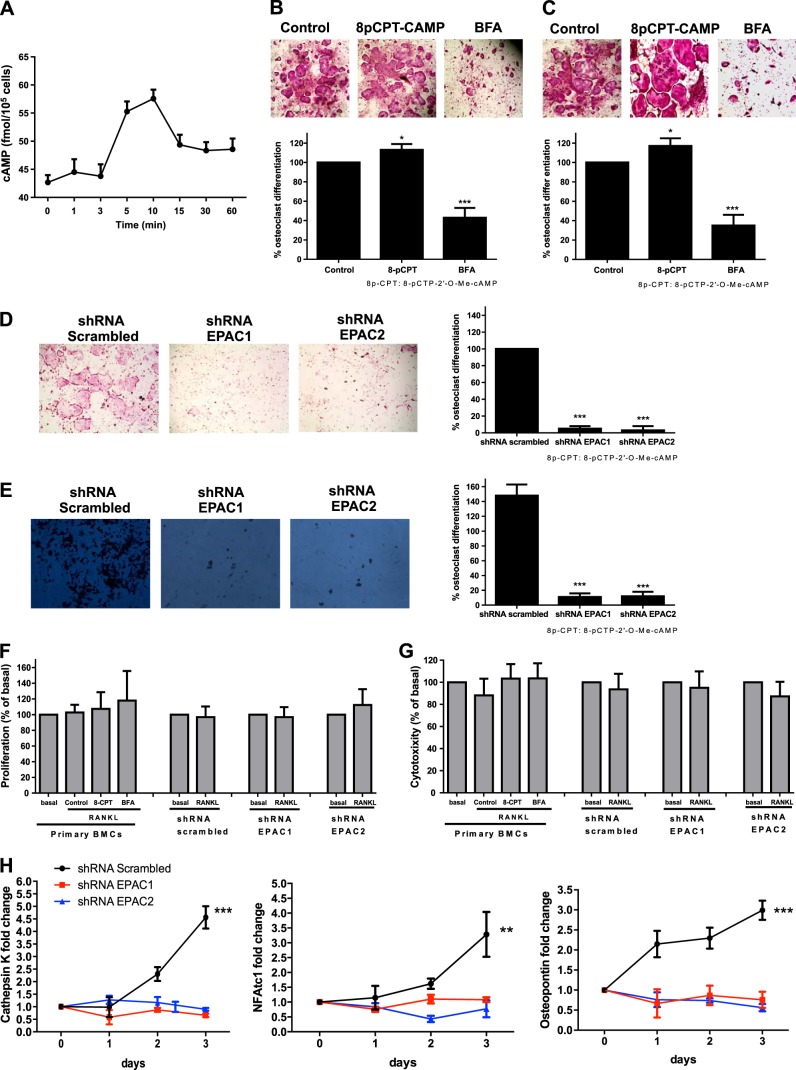
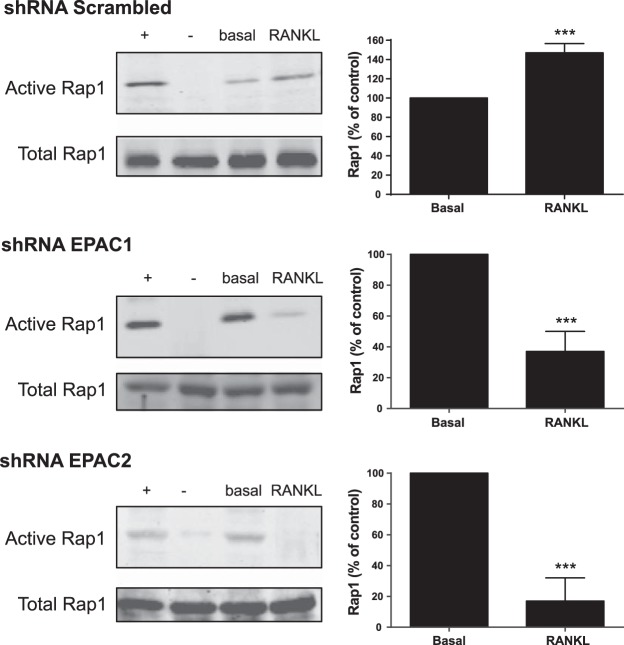
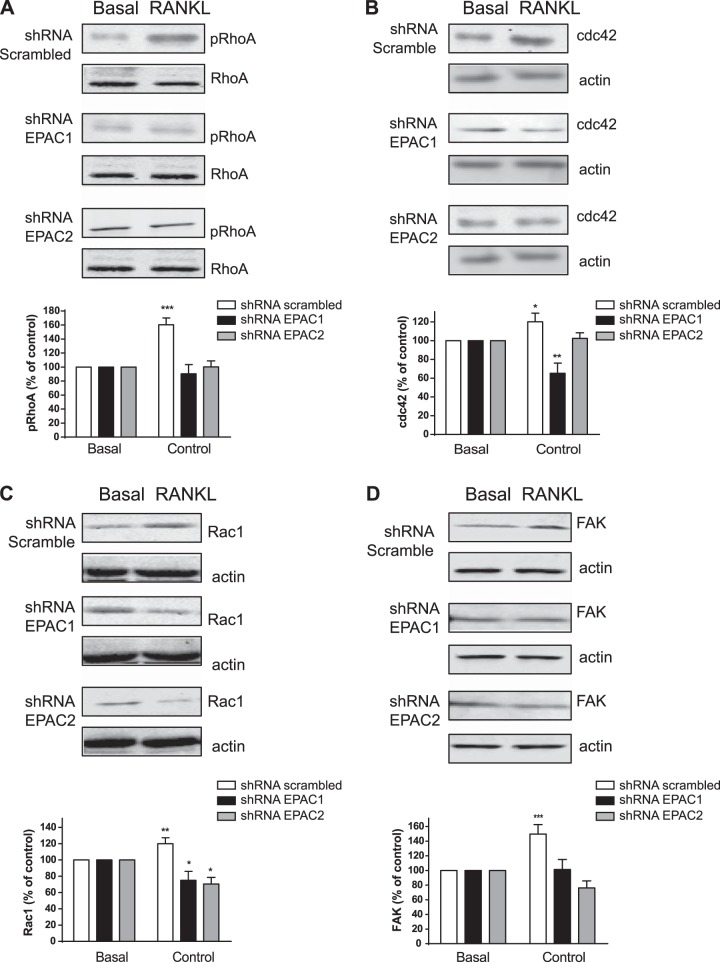
Bisphosphonates inhibit osteoclast differentiation/function via inhibition of Rap1A isoprenylation. As Rap1 is the effector of exchange protein directly activated by cAMP (EPAC) proteins, we determined the role of EPAC in osteoclast differentiation. We examined osteoclast differentiation as the number of primary murine/human bone-marrow precursors that differentiated into multinucleated TRAP-positive cells in the presence of EPAC-selective stimulus (8-pCTP-2'-O-Me-cAMP, 100 μM; 8-pCTP-2'-O-Me-cAMP-AM, 1 μM) or inhibitor brefeldin A (BFA), ESI-05, and ESI-09 (10 μM each). Rap1 activity was assessed, and signaling events, as well as differentiation in EPAC1/2-knockdown RAW264.7 cells, were studied. Direct EPAC1/2 stimulation significantly increased osteoclast differentiation, whereas EPAC1/2 inhibition diminished differentiation (113 ± 6%, P < 0.05, and 42 ± 10%, P < 0.001, of basal, respectively). Rap1 activation was maximal 15 min after RANKL stimulation (147 ± 9% of basal, P < 0.001), whereas silencing of EPAC1/2 diminished activated Rap1 (43 ± 13 and 20 ± 15% of control, P < 0.001) and NFkB nuclear translocation. TRAP-staining revealed no osteoclast differentiation in EPAC1/2-KO cells. Cathepsin K, NFATc1, and osteopontin mRNA expression decreased in EPAC1/2-KO cells when compared to control. RhoA, cdc42, Rac1, and FAK were activated in an EPAC1/2-dependent manner, and there was diminished cytoskeletal assembly in EPAC1/2-KO cells. In summary, EPAC1 and EPAC2 are critical signaling intermediates in osteoclast differentiation that permit RANKL-stimulated NFkB nuclear translocation and actin rearrangements. Targeting this signaling intermediate may diminish bone destruction in inflammatory arthritis.
Keywords: FAK; Rac1; RhoA; cdc42.
© FASEB.
Figures



References
-
- Suda T., Takahashi N., Udagawa N., Jimi E., Gillespie M. T., Martin T. J. (1999) Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 20, 345–357 - PubMed
-
- Manolagas S. C. (2000) Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr. Rev. 21, 115–137 - PubMed
-
- Teitelbaum S. L. (2000) Bone resorption by osteoclasts. Science 289, 1504–1508 - PubMed
-
- McHugh K. P., Shen Z., Crotti T. N., Flannery M. R., Fajardo R., Bierbaum B. E., Goldring S. R. (2007) Role of cell-matrix interactions in osteoclast differentiation. Adv. Exp. Med. Biol. 602, 107–111 - PubMed
-
- Takayanagi H. (2007) Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 7, 292–304 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
