

Complexin inhibits spontaneous release and synchronizes Ca2+-triggered synaptic vesicle fusion by distinct mechanisms
- PMID: 25122624
- PMCID: PMC4130161
- DOI: 10.7554/eLife.03756
Complexin inhibits spontaneous release and synchronizes Ca2+-triggered synaptic vesicle fusion by distinct mechanisms
Abstract
Previously we showed that fast Ca(2+)-triggered vesicle fusion with reconstituted neuronal SNAREs and synaptotagmin-1 begins from an initial hemifusion-free membrane point contact, rather than a hemifusion diaphragm, using a single vesicle-vesicle lipid/content mixing assay (Diao et al., 2012). When complexin-1 was included, a more pronounced Ca(2+)-triggered fusion burst was observed, effectively synchronizing the process. Here we show that complexin-1 also reduces spontaneous fusion in the same assay. Moreover, distinct effects of several complexin-1 truncation mutants on spontaneous and Ca(2+)-triggered fusion closely mimic those observed in neuronal cultures. The very N-terminal domain is essential for synchronization of Ca(2+)-triggered fusion, but not for suppression of spontaneous fusion, whereas the opposite is true for the C-terminal domain. By systematically varying the complexin-1 concentration, we observed differences in titration behavior for spontaneous and Ca(2+)-triggered fusion. Taken together, complexin-1 utilizes distinct mechanisms for synchronization of Ca(2+)-triggered fusion and inhibition of spontaneous fusion.
Keywords: SNARE; complexin; membrane fusion; neurotransmitter release; synaptic vesicle fusion; synaptotagmin.
Copyright © 2014, Lai et al.
Conflict of interest statement
ATB: Reviewing editor,
The other authors declare that no competing interests exist.
Figures
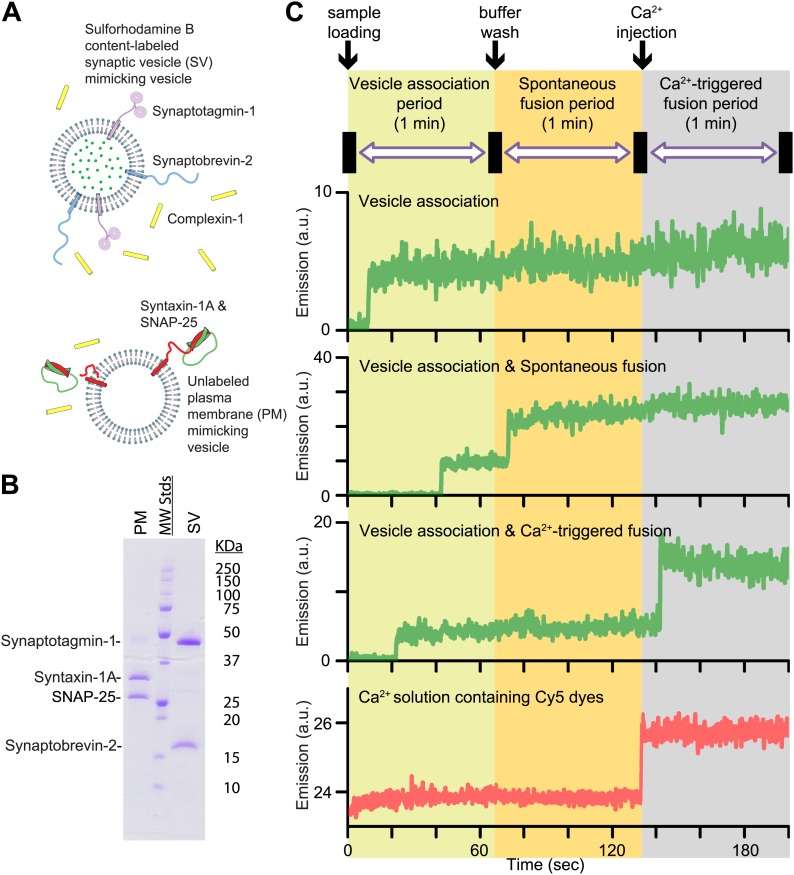
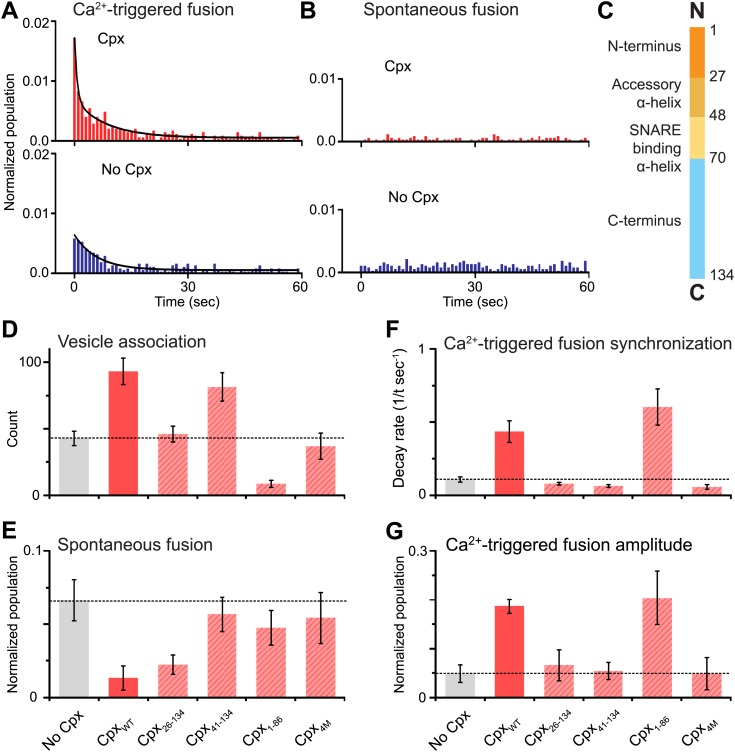
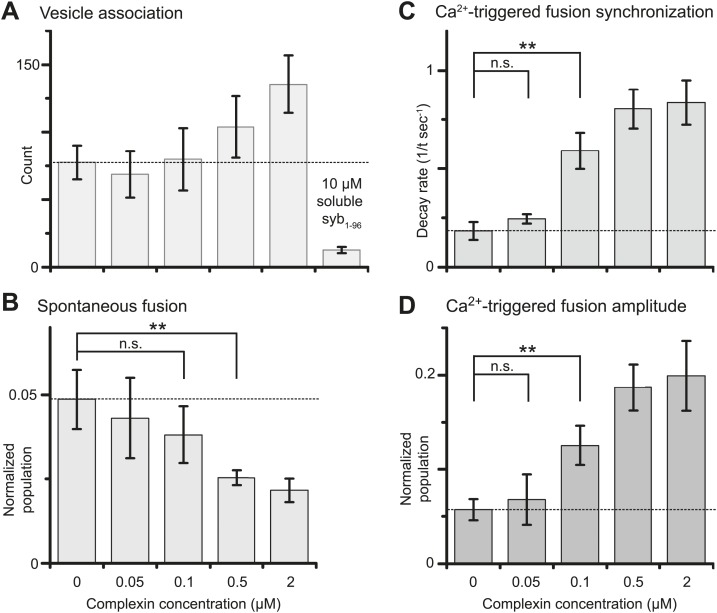
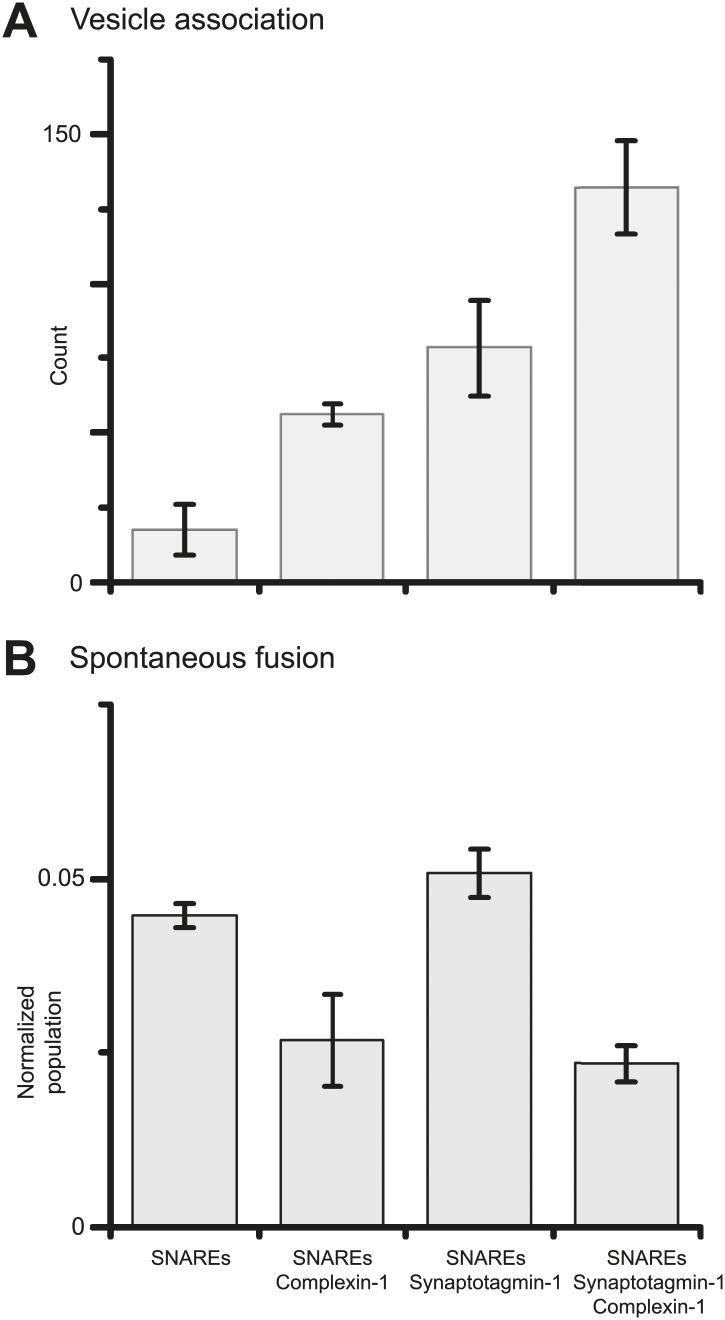
References
-
- Diao J, Grob P, Cipriano DJ, Kyoung M, Zhang Y, Shah S, Nguyen A, Padolina M, Srivastava A, Vrljic M, Shah A, Nogales E, Chu S, Brunger AT. 2012. Synaptic proteins promote calcium-triggered fast transition from point contact to full fusion. eLife 1:e00109. doi: 10.7554/eLife.00109 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
