

Multisensory training improves auditory spatial processing following bilateral cochlear implantation
- PMID: 25122908
- PMCID: PMC4131019
- DOI: 10.1523/JNEUROSCI.4767-13.2014
Multisensory training improves auditory spatial processing following bilateral cochlear implantation
Abstract
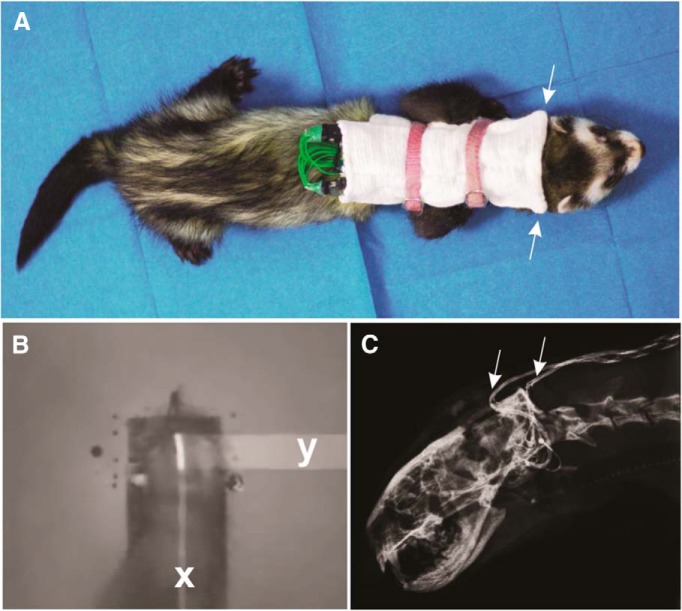
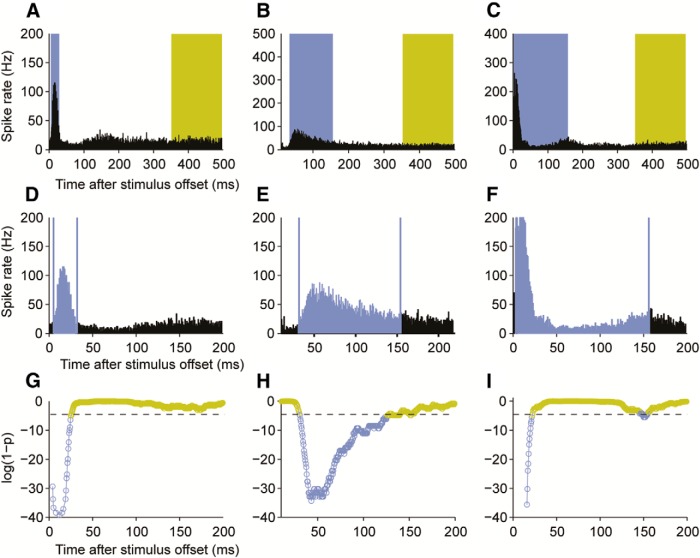
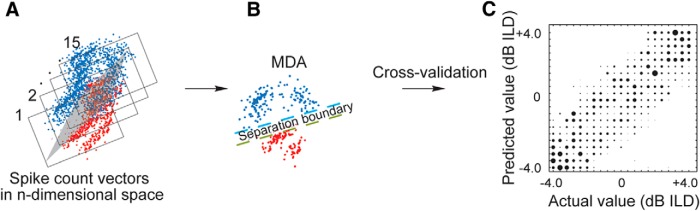
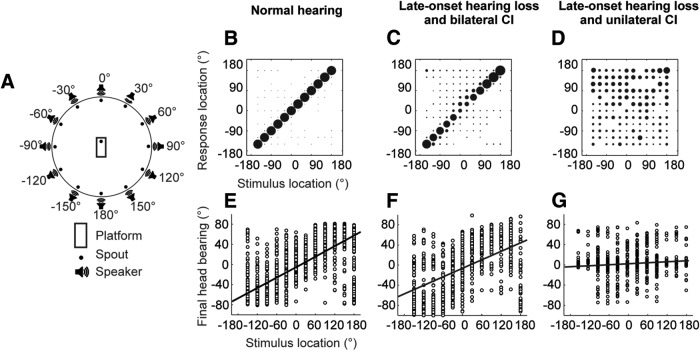
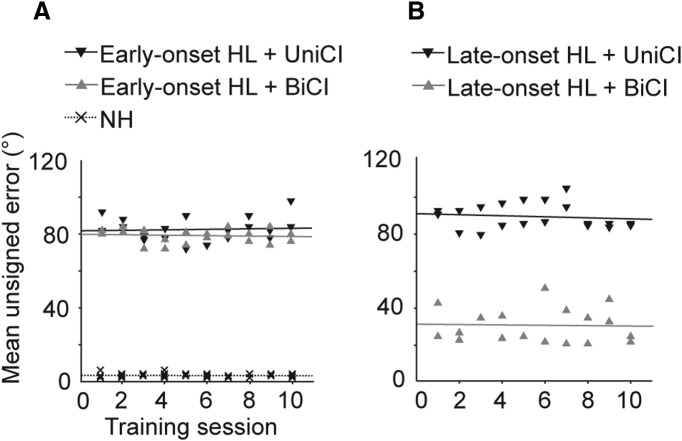
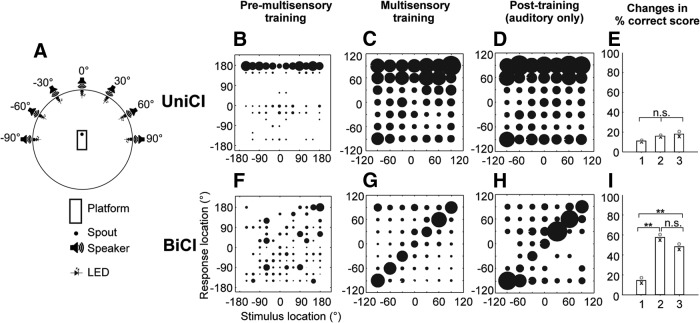
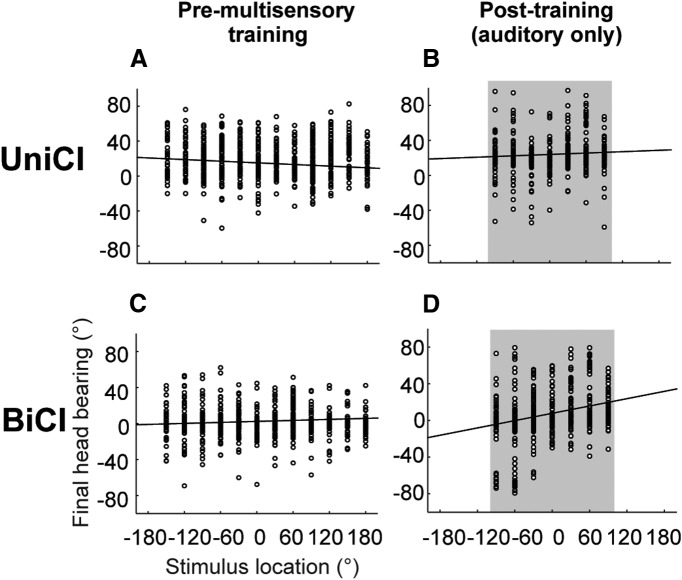
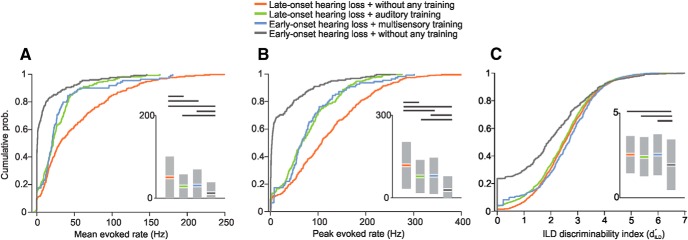
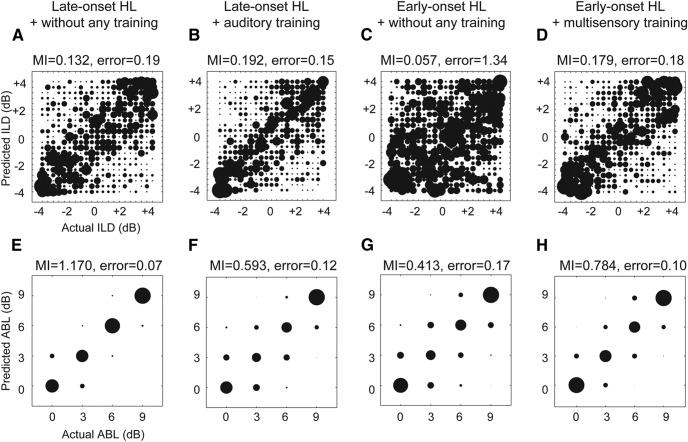
Cochlear implants (CIs) partially restore hearing to the deaf by directly stimulating the inner ear. In individuals fitted with CIs, lack of auditory experience due to loss of hearing before language acquisition can adversely impact outcomes. For example, adults with early-onset hearing loss generally do not integrate inputs from both ears effectively when fitted with bilateral CIs (BiCIs). Here, we used an animal model to investigate the effects of long-term deafness on auditory localization with BiCIs and approaches for promoting the use of binaural spatial cues. Ferrets were deafened either at the age of hearing onset or as adults. All animals were implanted in adulthood, either unilaterally or bilaterally, and were subsequently assessed for their ability to localize sound in the horizontal plane. The unilaterally implanted animals were unable to perform this task, regardless of the duration of deafness. Among animals with BiCIs, early-onset hearing loss was associated with poor auditory localization performance, compared with late-onset hearing loss. However, performance in the early-deafened group with BiCIs improved significantly after multisensory training with interleaved auditory and visual stimuli. We demonstrate a possible neural substrate for this by showing a training-induced improvement in the responsiveness of auditory cortical neurons and in their sensitivity to interaural level differences, the principal localization cue available to BiCI users. Importantly, our behavioral and physiological evidence demonstrates a facilitative role for vision in restoring auditory spatial processing following potential cross-modal reorganization. These findings support investigation of a similar training paradigm in human CI users.
Keywords: auditory cortex; cochlear implant; cross-modal plasticity; hearing loss; multisensory; sound localization.
Copyright © 2014 Isaiah et al.
Figures
Similar articles
-
Sound source localization patterns and bilateral cochlear implants: Age at onset of deafness effects.PLoS One. 2022 Feb 8;17(2):e0263516. doi: 10.1371/journal.pone.0263516. eCollection 2022. PLoS One. 2022. PMID: 35134072 Free PMC article.
-
Cortical Representation of Interaural Time Difference Is Impaired by Deafness in Development: Evidence from Children with Early Long-term Access to Sound through Bilateral Cochlear Implants Provided Simultaneously.J Neurosci. 2017 Mar 1;37(9):2349-2361. doi: 10.1523/JNEUROSCI.2538-16.2017. Epub 2017 Jan 25. J Neurosci. 2017. PMID: 28123078 Free PMC article.
-
Cortical Processing of Level Cues for Spatial Hearing is Impaired in Children with Prelingual Deafness Despite Early Bilateral Access to Sound.Brain Topogr. 2018 Mar;31(2):270-287. doi: 10.1007/s10548-017-0596-5. Epub 2017 Nov 8. Brain Topogr. 2018. PMID: 29119311
-
Bilateral cochlear implants in children: Effects of auditory experience and deprivation on auditory perception.Hear Res. 2016 Aug;338:76-87. doi: 10.1016/j.heares.2016.01.003. Epub 2016 Jan 30. Hear Res. 2016. PMID: 26828740 Free PMC article. Review.
-
[Effect of cochlear implantation on sound localization for patients with unilateral sensorineural hearing loss].Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2016 Aug 7;51(8):623-30. doi: 10.3760/cma.j.issn.1673-0860.2016.08.015. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2016. PMID: 27625137 Review. Chinese.
Cited by
-
Auditory cortical plasticity in cochlear implant users.Curr Opin Neurobiol. 2020 Feb;60:108-114. doi: 10.1016/j.conb.2019.11.003. Epub 2019 Dec 18. Curr Opin Neurobiol. 2020. PMID: 31864104 Free PMC article. Review.
-
Re-weighting of Sound Localization Cues by Audiovisual Training.Front Neurosci. 2019 Nov 15;13:1164. doi: 10.3389/fnins.2019.01164. eCollection 2019. Front Neurosci. 2019. PMID: 31802997 Free PMC article.
-
Having Two Ears Facilitates the Perceptual Separation of Concurrent Talkers for Bilateral and Single-Sided Deaf Cochlear Implantees.Ear Hear. 2016 May-Jun;37(3):289-302. doi: 10.1097/AUD.0000000000000284. Ear Hear. 2016. PMID: 26886027 Free PMC article.
-
Behavioural benefits of multisensory processing in ferrets.Eur J Neurosci. 2017 Jan;45(2):278-289. doi: 10.1111/ejn.13440. Epub 2016 Nov 3. Eur J Neurosci. 2017. PMID: 27740711 Free PMC article.
-
Cross-Modal Plasticity in Higher-Order Auditory Cortex of Congenitally Deaf Cats Does Not Limit Auditory Responsiveness to Cochlear Implants.J Neurosci. 2016 Jun 8;36(23):6175-85. doi: 10.1523/JNEUROSCI.0046-16.2016. J Neurosci. 2016. PMID: 27277796 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical