

Daily torpor and hibernation in birds and mammals
- PMID: 25123049
- PMCID: PMC4351926
- DOI: 10.1111/brv.12137
Daily torpor and hibernation in birds and mammals
Abstract
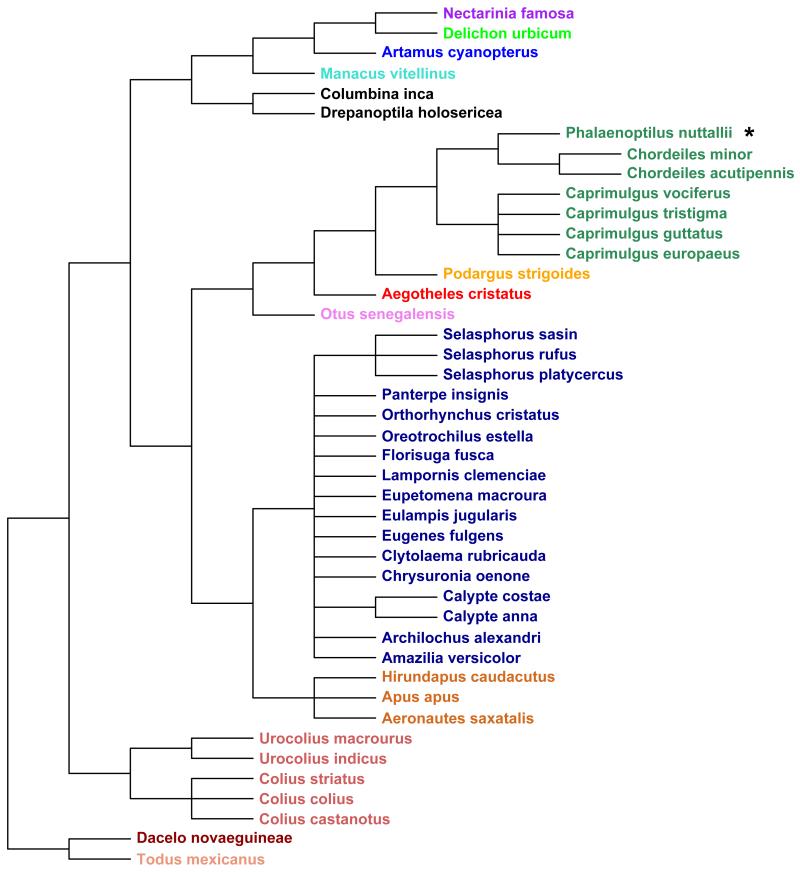
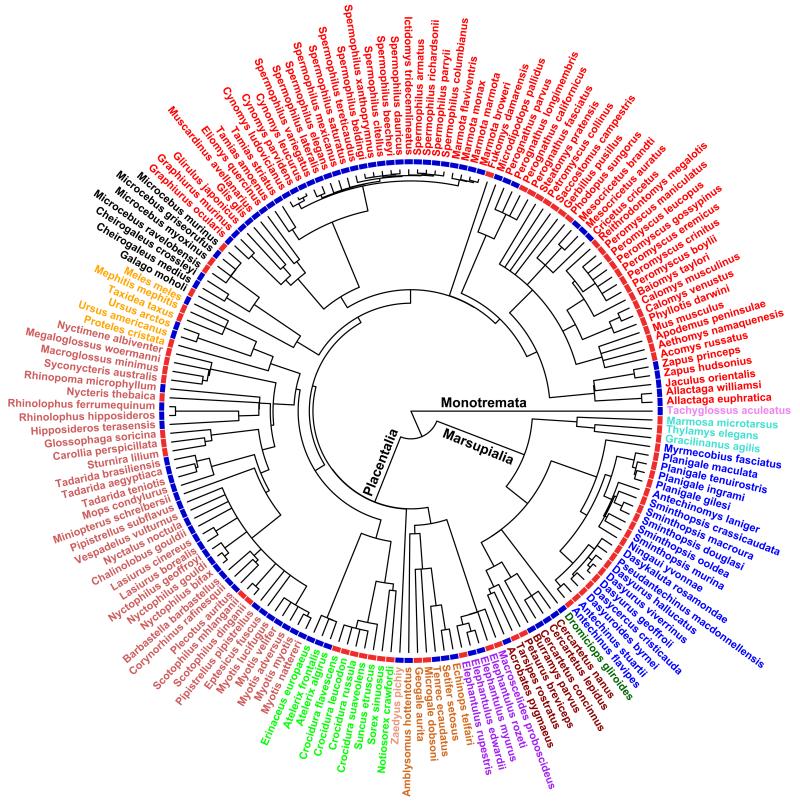
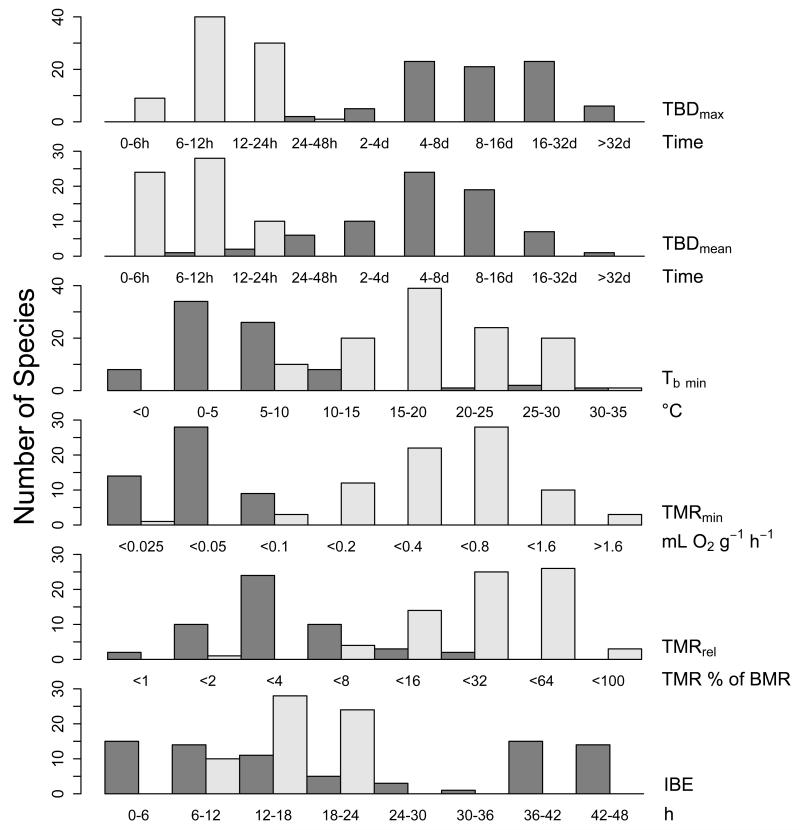
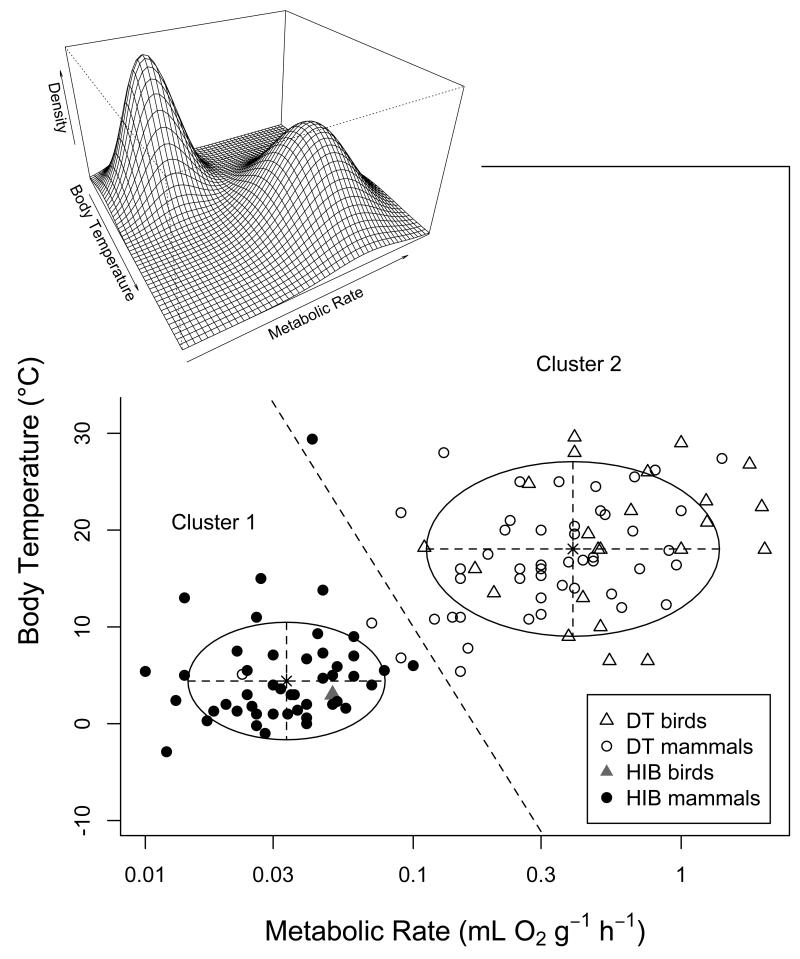
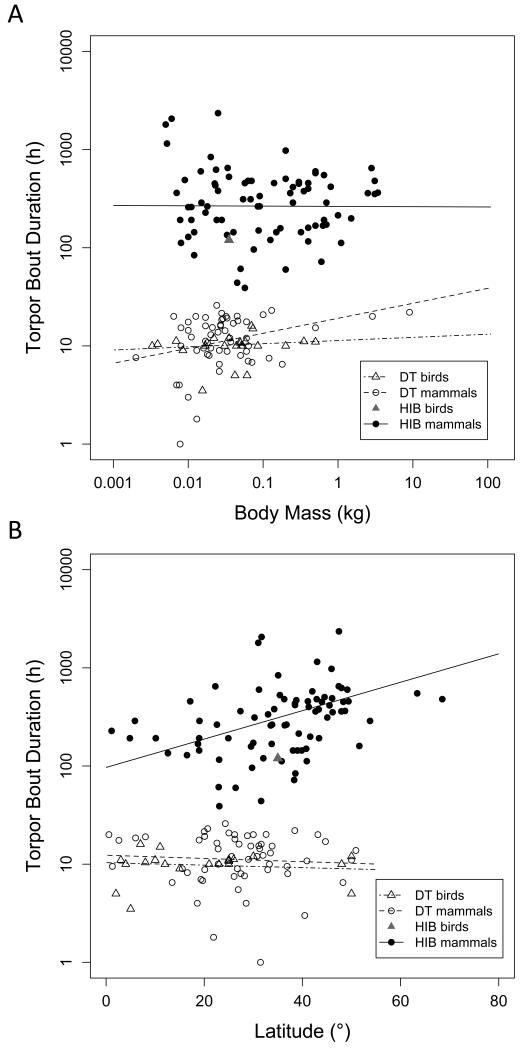
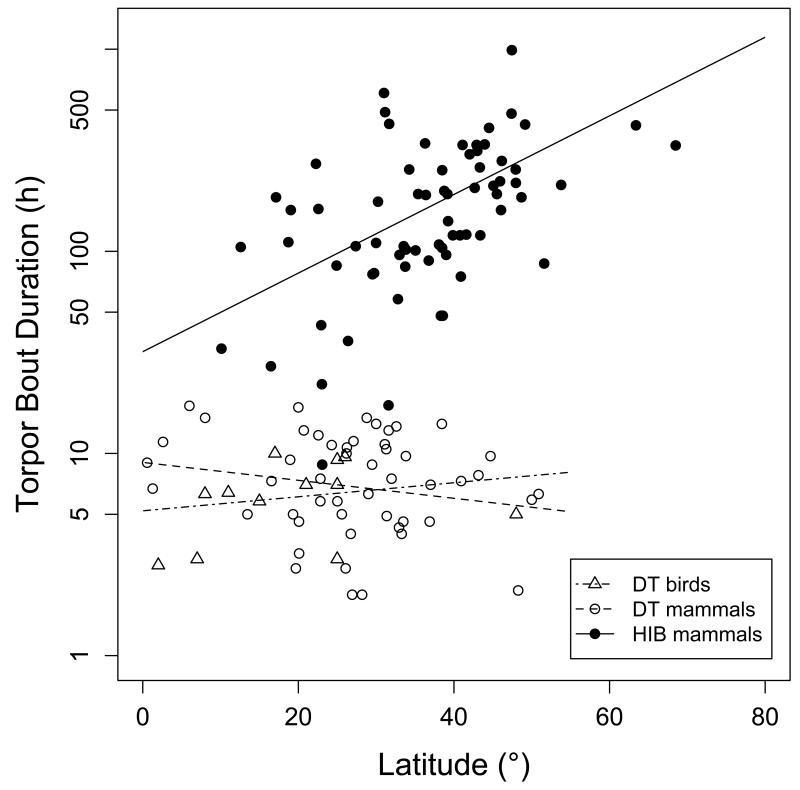
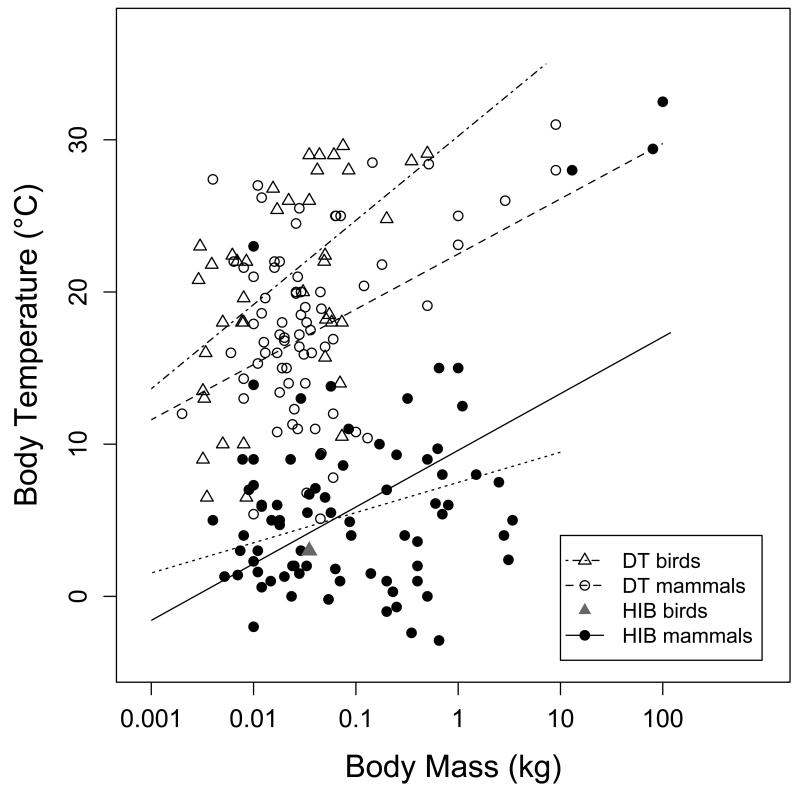
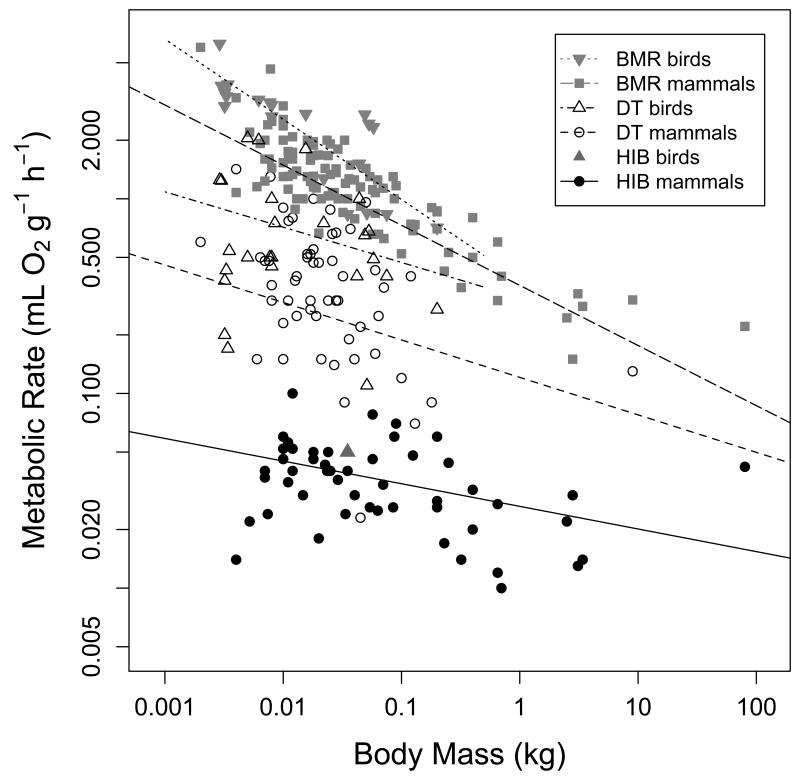
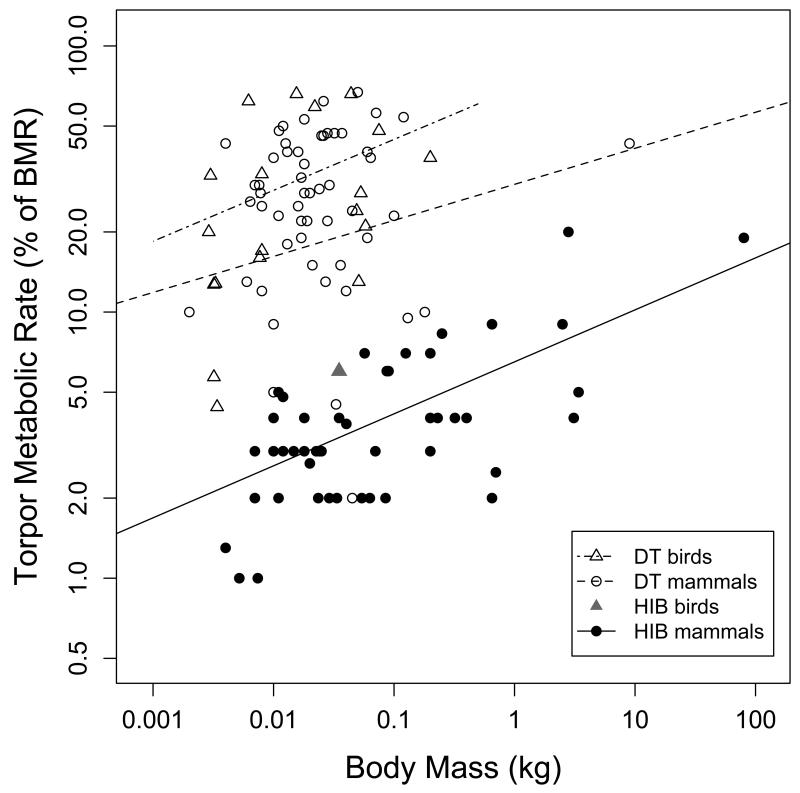
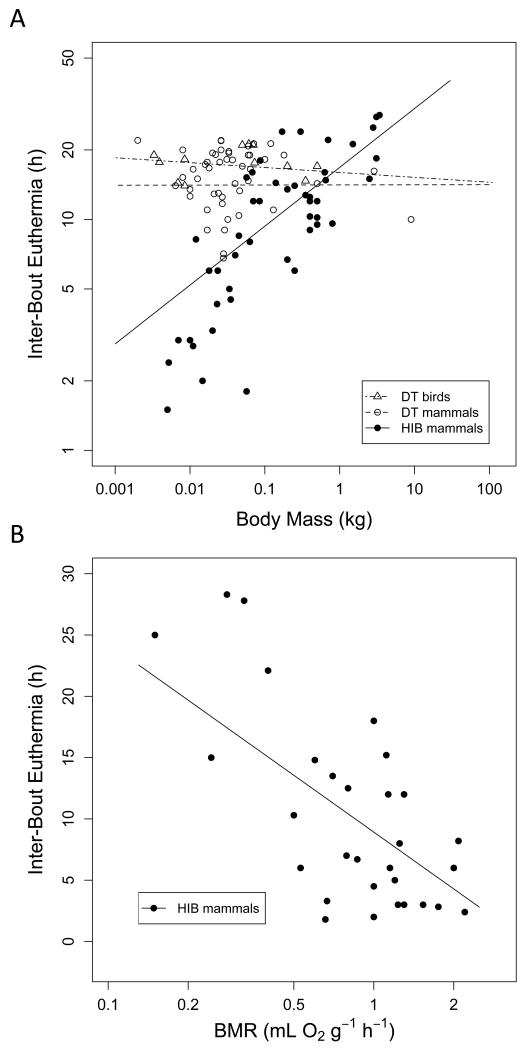
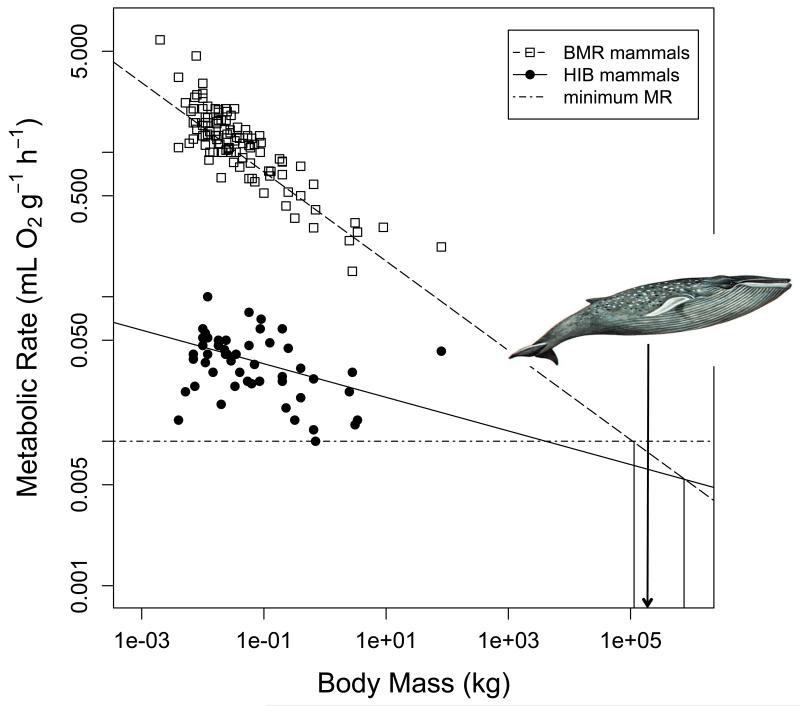
Many birds and mammals drastically reduce their energy expenditure during times of cold exposure, food shortage, or drought, by temporarily abandoning euthermia, i.e. the maintenance of high body temperatures. Traditionally, two different types of heterothermy, i.e. hypometabolic states associated with low body temperature (torpor), have been distinguished: daily torpor, which lasts less than 24 h and is accompanied by continued foraging, versus hibernation, with torpor bouts lasting consecutive days to several weeks in animals that usually do not forage but rely on energy stores, either food caches or body energy reserves. This classification of torpor types has been challenged, suggesting that these phenotypes may merely represent extremes in a continuum of traits. Here, we investigate whether variables of torpor in 214 species (43 birds and 171 mammals) form a continuum or a bimodal distribution. We use Gaussian-mixture cluster analysis as well as phylogenetically informed regressions to quantitatively assess the distinction between hibernation and daily torpor and to evaluate the impact of body mass and geographical distribution of species on torpor traits. Cluster analysis clearly confirmed the classical distinction between daily torpor and hibernation. Overall, heterothermic endotherms tend to be small; hibernators are significantly heavier than daily heterotherms and also are distributed at higher average latitudes (∼35°) than daily heterotherms (∼25°). Variables of torpor for an average 30 g heterotherm differed significantly between daily heterotherms and hibernators. Average maximum torpor bout duration was >30-fold longer, and mean torpor bout duration >25-fold longer in hibernators. Mean minimum body temperature differed by ∼13°C, and the mean minimum torpor metabolic rate was ∼35% of the basal metabolic rate (BMR) in daily heterotherms but only 6% of BMR in hibernators. Consequently, our analysis strongly supports the view that hibernators and daily heterotherms are functionally distinct groups that probably have been subject to disruptive selection. Arguably, the primary physiological difference between daily torpor and hibernation, which leads to a variety of derived further distinct characteristics, is the temporal control of entry into and arousal from torpor, which is governed by the circadian clock in daily heterotherms, but apparently not in hibernators.
Keywords: daily torpor; endotherms; energy savings; heterothermy; hibernation; hypometabolism; hypothermia; over‐wintering; thermoregulation.
© 2014 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
References
-
- Anderson MD. Aardwolf adaptations: a review. Transactions of the Royal Society of South Africa. 2004;59:99–104. DOI: 10.1080/00359190409519168.
-
- Arlettaz R, Ruchet C, Aeschimann J, Brun E, Genoud M, Vogel P. Physiological traits affecting the distribution and wintering strategy of the bat Tadarida teniotis. Ecology. 2000;81:1004–1014. DOI: 10.1890/0012-9658(2000)081[1004:PTATDA]2.0.CO;2.
-
- Armitage KB, Woods BC, Salsbury CM. In: Heldmaier G, Klingenspor M, editors. Energetics of hibernation in woodchucks (Marmota monax); Life in the Cold. 11th International Hibernation Symposium; Springer, Berlin Heidelberg New York. 2000.pp. 73–80.
-
- Arnold JM. Dasyurus geoffroii. University of Western Australia; 1976. Growth and bioenergetics of the Chuditch.
-
- Arnold W. Energetics of social hibernation. In: Carey C, Florant GL, Wunder BA, Horwitz B, editors. Life in the Cold: Ecological, Physiological, and Molecular Mechanisms. Westview Press; Boulder: 1993. pp. 65–80.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
