

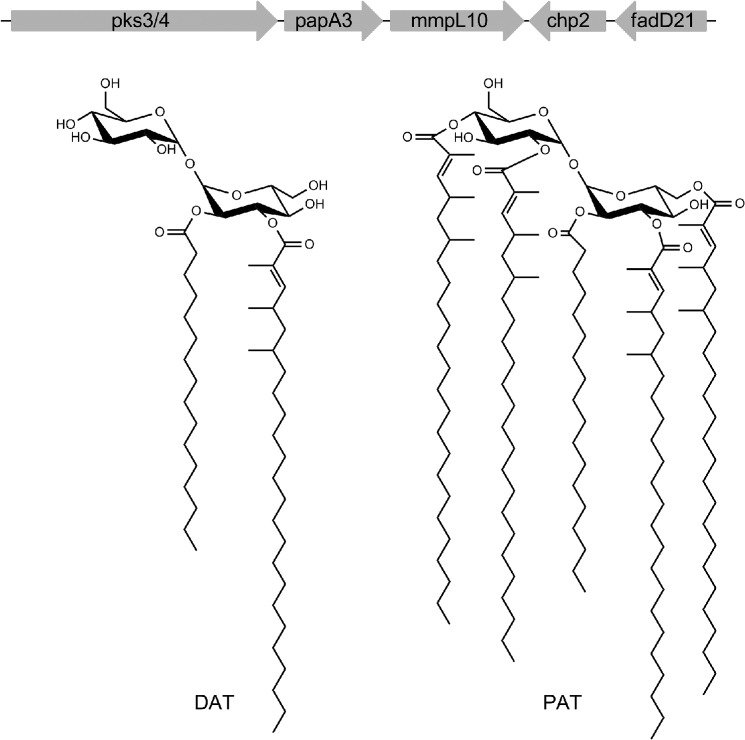
Biosynthesis and translocation of unsulfated acyltrehaloses in Mycobacterium tuberculosis
- PMID: 25124040
- PMCID: PMC4183827
- DOI: 10.1074/jbc.M114.581199
Biosynthesis and translocation of unsulfated acyltrehaloses in Mycobacterium tuberculosis
Abstract
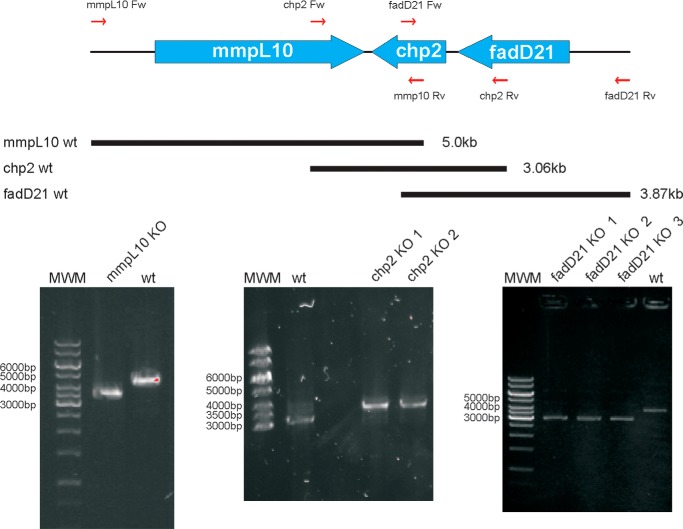
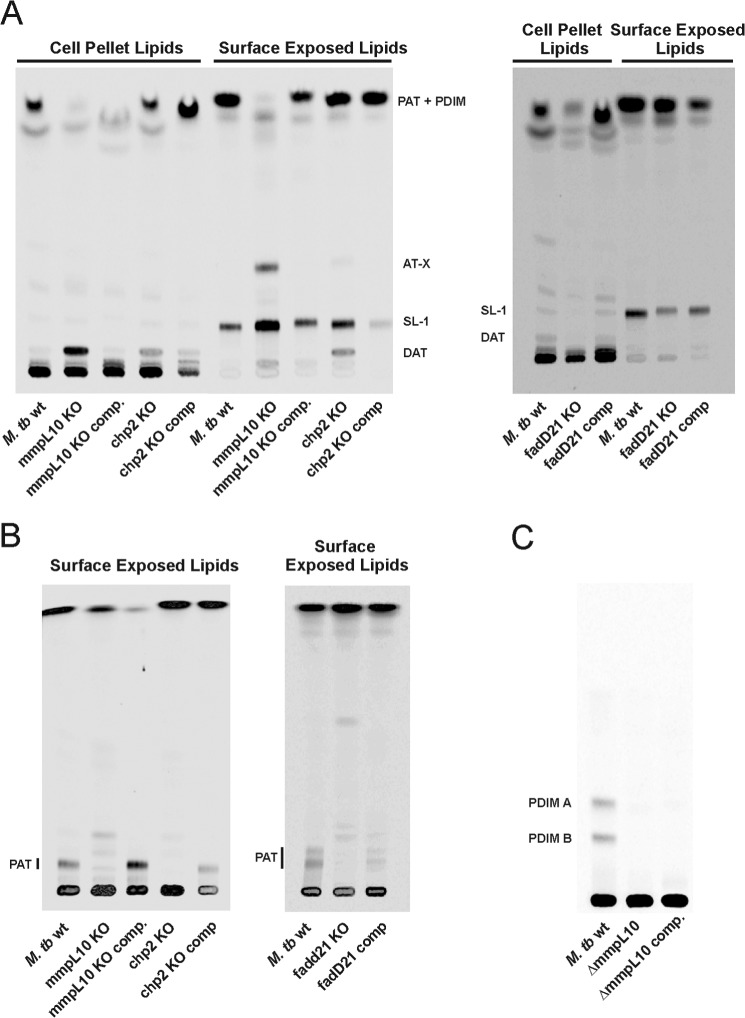
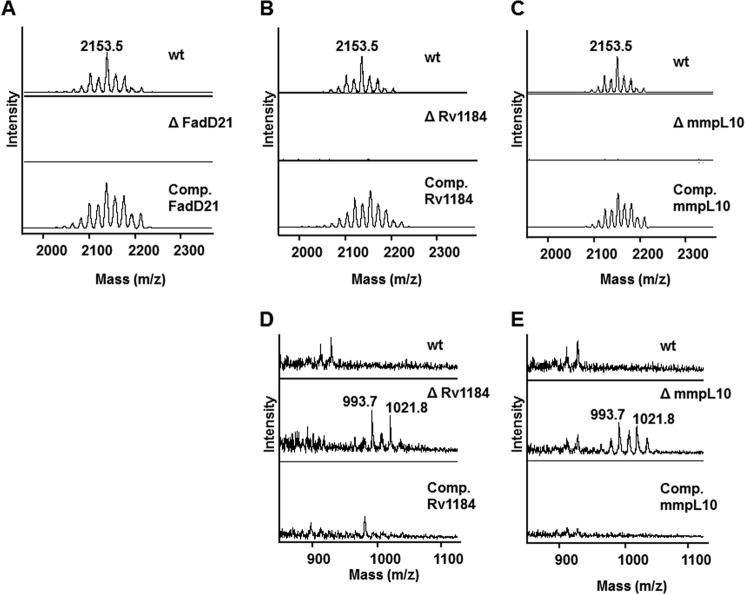
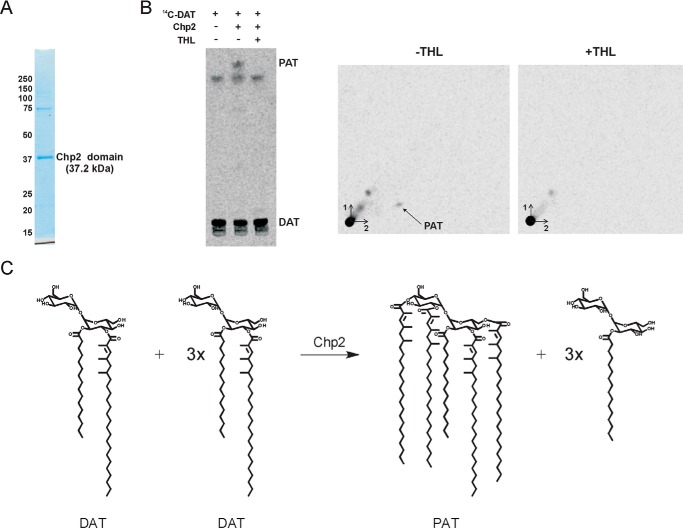
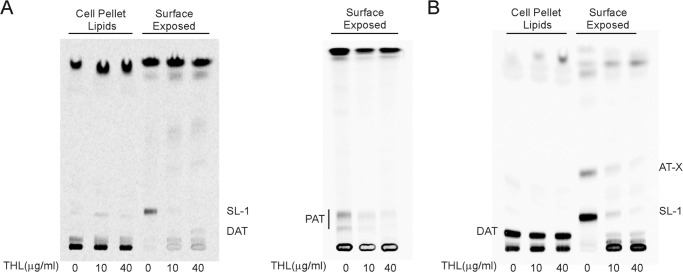
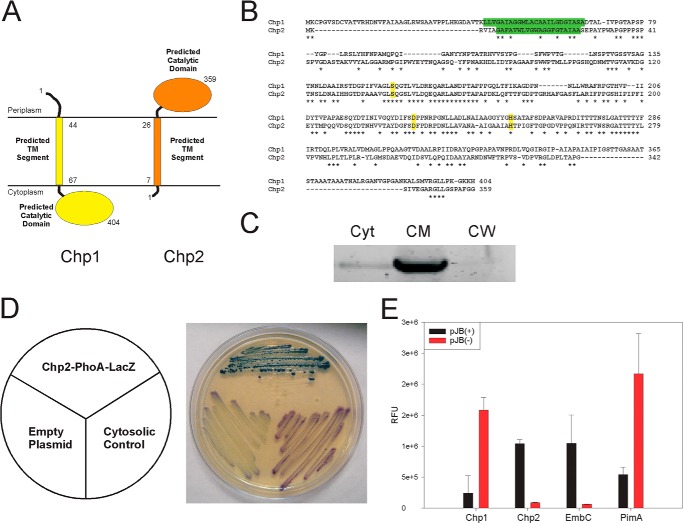
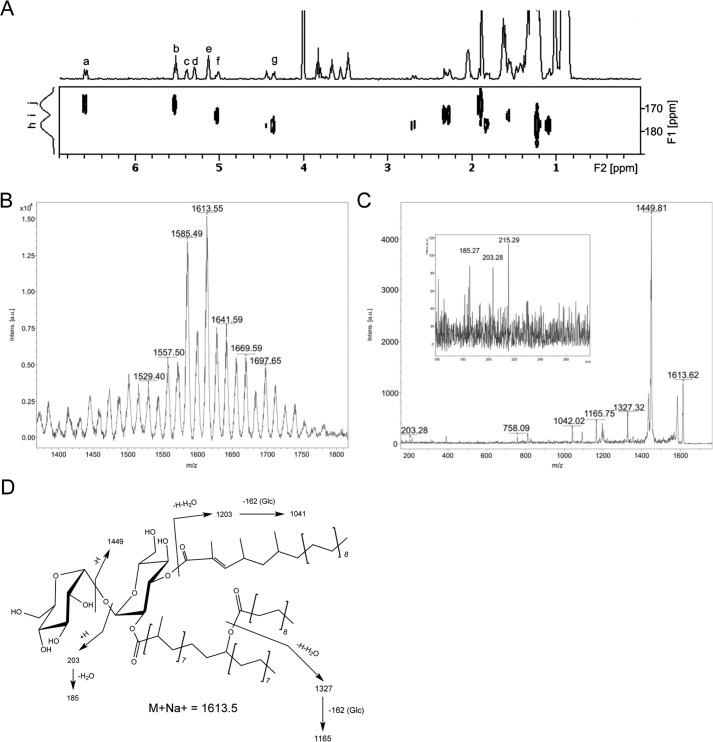
A number of species-specific polymethyl-branched fatty acid-containing trehalose esters populate the outer membrane of Mycobacterium tuberculosis. Among them, 2,3-diacyltrehaloses (DAT) and penta-acyltrehaloses (PAT) not only play a structural role in the cell envelope but also contribute to the ability of M. tuberculosis to multiply and persist in the infected host, promoting the intracellular survival of the bacterium and modulating host immune responses. The nature of the machinery, topology, and sequential order of the reactions leading to the biosynthesis, assembly, and export of these complex glycolipids to the cell surface are the object of the present study. Our genetic and biochemical evidence corroborates a model wherein the biosynthesis and translocation of DAT and PAT to the periplasmic space are coupled and topologically split across the plasma membrane. The formation of DAT occurs on the cytosolic face of the plasma membrane through the action of PapA3, FadD21, and Pks3/4; that of PAT occurs on the periplasmic face via transesterification reactions between DAT substrates catalyzed by the acyltransferase Chp2 (Rv1184c). The integral membrane transporter MmpL10 is essential for DAT to reach the cell surface, and its presence in the membrane is required for Chp2 to be active. Disruption of mmpL10 or chp2 leads to an important build-up of DAT inside the cells and to the formation of a novel form of unsulfated acyltrehalose esterified with polymethyl-branched fatty acids normally found in sulfolipids that is translocated to the cell surface.
Keywords: Acyltransferase; Acyltrehalose; Chp2; Glycolipid; MmpL10; Mycobacteria; Polyketide; Transporter; Tuberculosis.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Daffé M., Draper P. (1998) The envelope layers of mycobacteria with reference to their pathogenicity. Adv. Microb. Physiol. 39, 131–203 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
