

State-dependent architecture of thalamic reticular subnetworks
- PMID: 25126786
- PMCID: PMC4205482
- DOI: 10.1016/j.cell.2014.06.025
State-dependent architecture of thalamic reticular subnetworks
Abstract
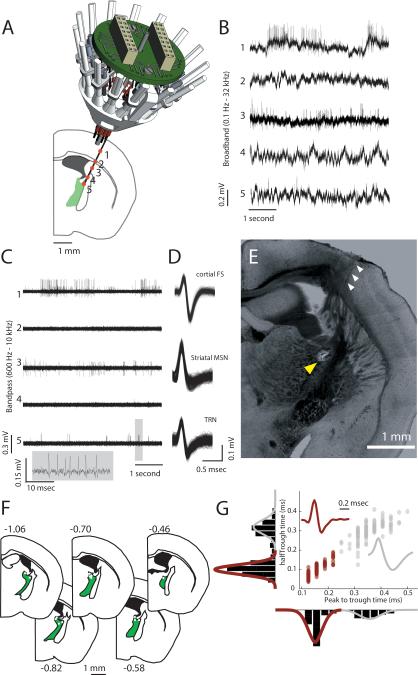
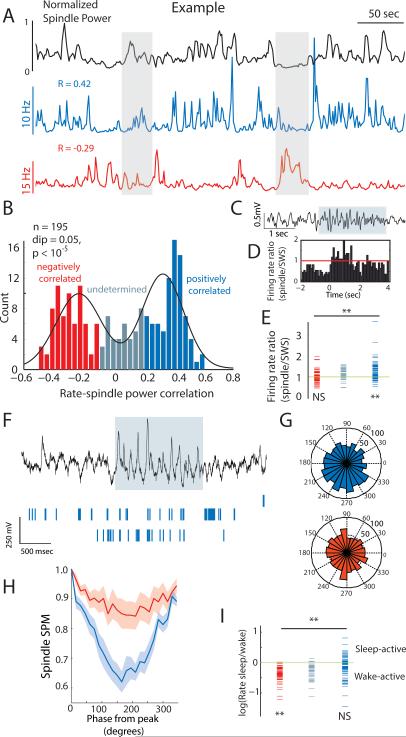
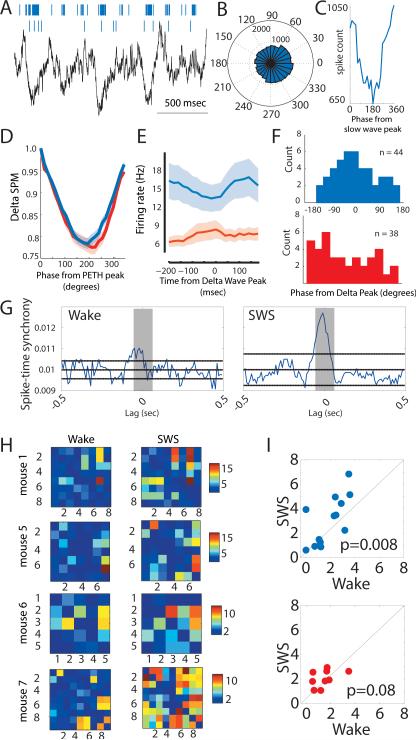
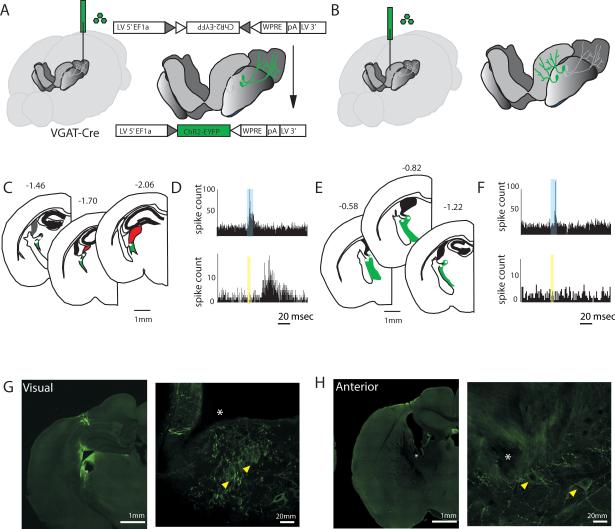
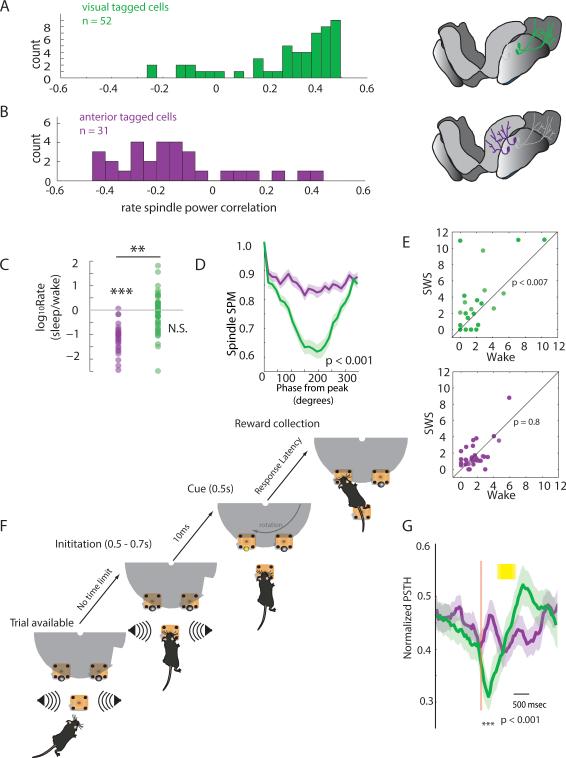
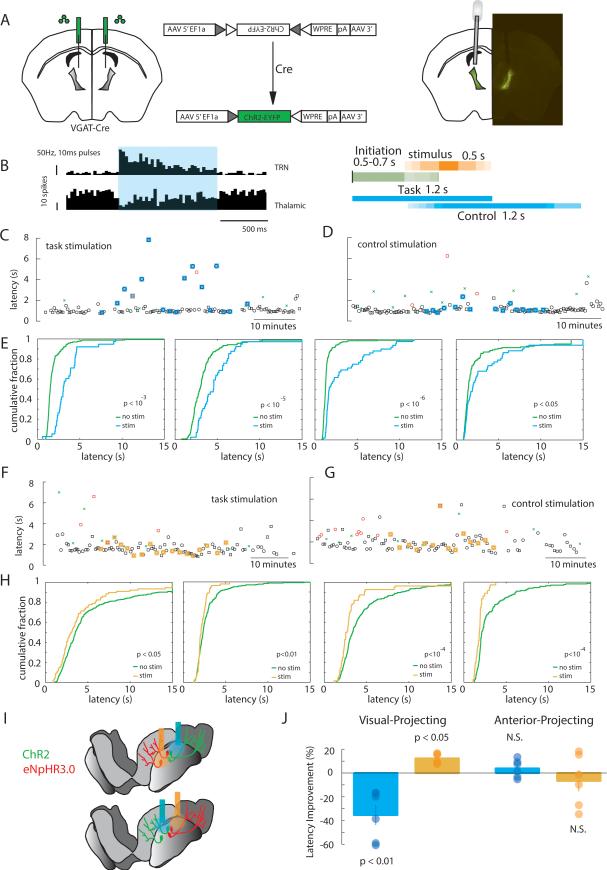
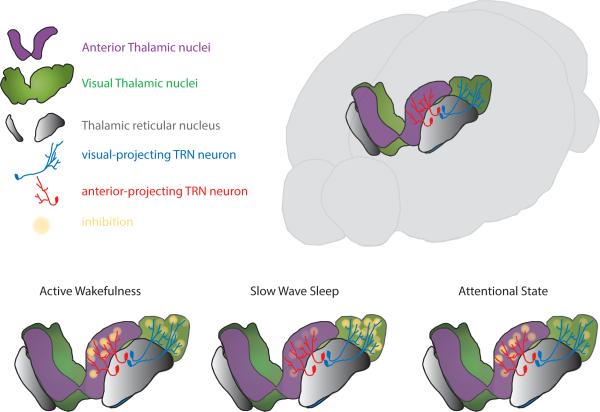
Behavioral state is known to influence interactions between thalamus and cortex, which are important for sensation, action, and cognition. The thalamic reticular nucleus (TRN) is hypothesized to regulate thalamo-cortical interactions, but the underlying functional architecture of this process and its state dependence are unknown. By combining the first TRN ensemble recording with psychophysics and connectivity-based optogenetic tagging, we found reticular circuits to be composed of distinct subnetworks. While activity of limbic-projecting TRN neurons positively correlates with arousal, sensory-projecting neurons participate in spindles and show elevated synchrony by slow waves during sleep. Sensory-projecting neurons are suppressed by attentional states, demonstrating that their gating of thalamo-cortical interactions is matched to behavioral state. Bidirectional manipulation of attentional performance was achieved through subnetwork-specific optogenetic stimulation. Together, our findings provide evidence for differential inhibition of thalamic nuclei across brain states, where the TRN separately controls external sensory and internal limbic processing facilitating normal cognitive function. PAPERFLICK:
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Bargmann CI, Marder E. From the connectome to brain function. Nat Methods. 2013;10:483–490. - PubMed
-
- Bazhenov M, Timofeev I, Steriade M, Sejnowski T. Spiking-bursting activity in the thalamic reticular nucleus initiates sequences of spindle oscillations in thalamic networks. J Neurophysiol. 2000;84:1076–1087. - PubMed
-
- Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci. 2005;8:1263–1268. - PubMed
Publication types
MeSH terms
Grants and funding
- R01 MH061976/MH/NIMH NIH HHS/United States
- DP1MH103908/DP/NCCDPHP CDC HHS/United States
- R25 MH094612/MH/NIMH NIH HHS/United States
- K99 NS078115/NS/NINDS NIH HHS/United States
- R01-MH061976/MH/NIMH NIH HHS/United States
- R01NS077986/NS/NINDS NIH HHS/United States
- R01MH057414/MH/NIMH NIH HHS/United States
- R01 MH057414/MH/NIMH NIH HHS/United States
- K99 NS 078115/NS/NINDS NIH HHS/United States
- R01 NS077986/NS/NINDS NIH HHS/United States
- R01 MH101209/MH/NIMH NIH HHS/United States
- R01 GM104948/GM/NIGMS NIH HHS/United States
- DP1 MH103908/MH/NIMH NIH HHS/United States
- TR01-GM10498/GM/NIGMS NIH HHS/United States
- R01MH101209/MH/NIMH NIH HHS/United States
- R00 NS078115/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
