

Dissecting engineered cell types and enhancing cell fate conversion via CellNet
- PMID: 25126792
- PMCID: PMC4291075
- DOI: 10.1016/j.cell.2014.07.021
Dissecting engineered cell types and enhancing cell fate conversion via CellNet
Abstract
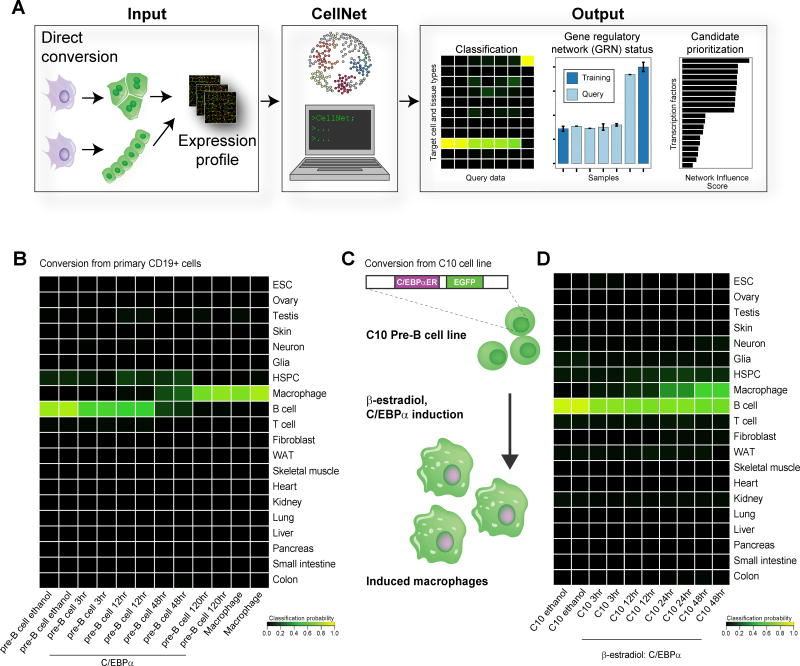
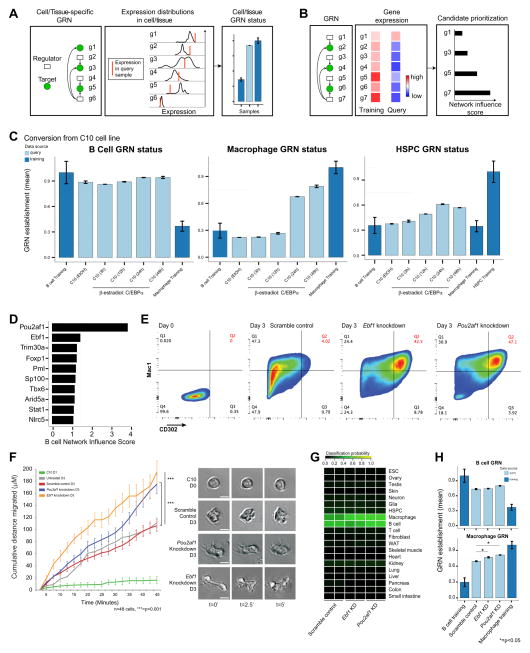
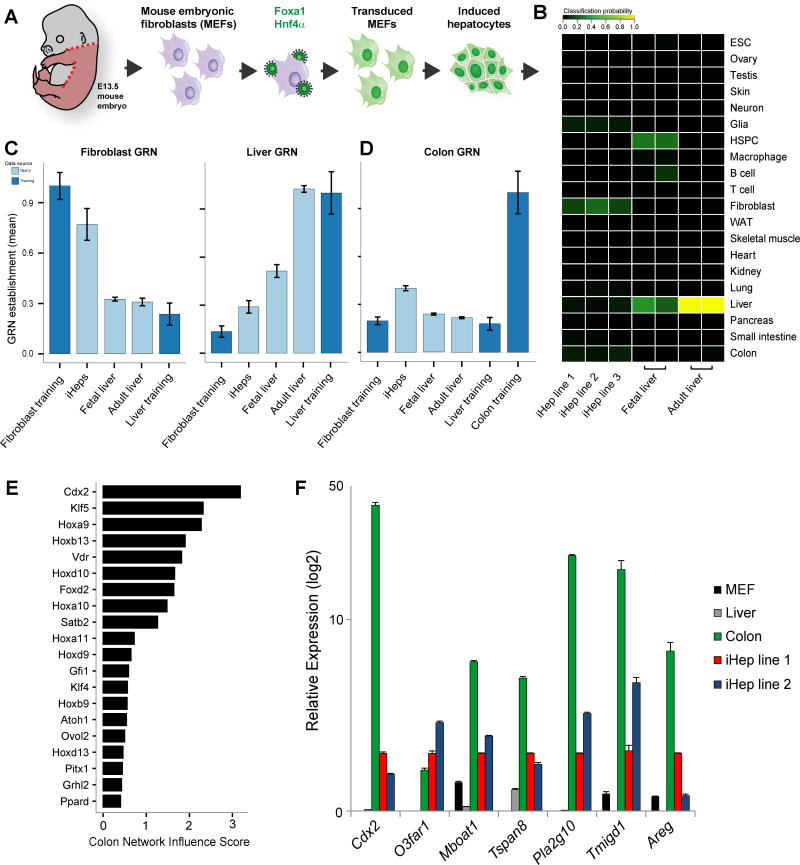
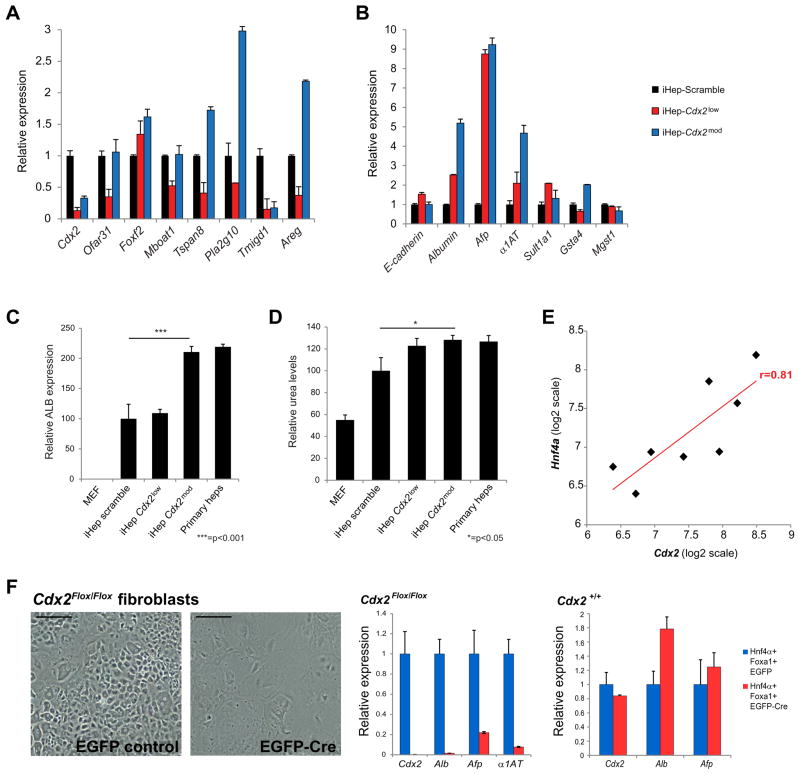
Engineering clinically relevant cells in vitro holds promise for regenerative medicine, but most protocols fail to faithfully recapitulate target cell properties. To address this, we developed CellNet, a network biology platform that determines whether engineered cells are equivalent to their target tissues, diagnoses aberrant gene regulatory networks, and prioritizes candidate transcriptional regulators to enhance engineered conversions. Using CellNet, we improved B cell to macrophage conversion, transcriptionally and functionally, by knocking down predicted B cell regulators. Analyzing conversion of fibroblasts to induced hepatocytes (iHeps), CellNet revealed an unexpected intestinal program regulated by the master regulator Cdx2. We observed long-term functional engraftment of mouse colon by iHeps, thereby establishing their broader potential as endoderm progenitors and demonstrating direct conversion of fibroblasts into intestinal epithelium. Our studies illustrate how CellNet can be employed to improve direct conversion and to uncover unappreciated properties of engineered cells.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
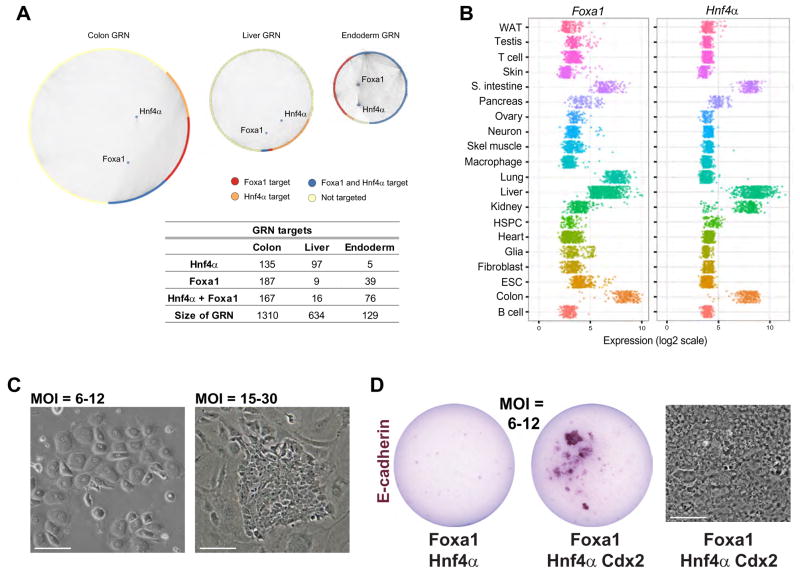
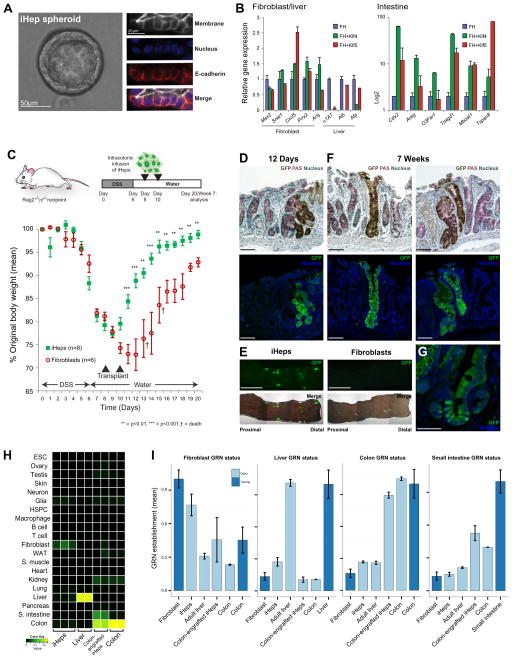
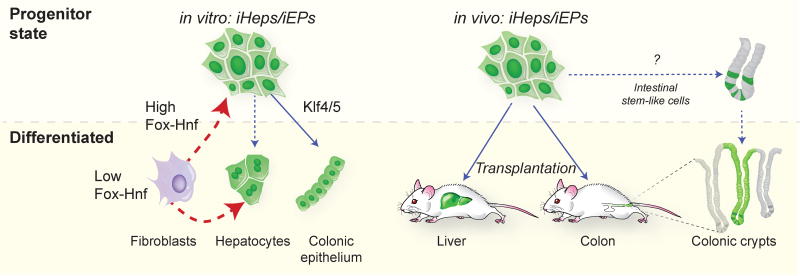
Comment in
-
Defining cellular identity through network biology.Cell Cycle. 2014;13(21):3313-4. doi: 10.4161/15384101.2014.972918. Cell Cycle. 2014. PMID: 25485572 Free PMC article. No abstract available.
References
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
-
- Bussmann LH, Schubert A, Vu Manh TP, De Andres L, Desbordes SC, Parra M, Zimmermann T, Rapino F, Rodriguez-Ubreva J, Ballestar E, et al. A robust and highly efficient immune cell reprogramming system. Cell Stem Cell. 2009;5:554–566. - PubMed
-
- Cohen DE, Melton D. Turning straw into gold: directing cell fate for regenerative medicine. Nat Rev Genet. 2011;12:243–252. - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
- T32 HL066987/HL/NHLBI NIH HHS/United States
- R24DK092760/DK/NIDDK NIH HHS/United States
- R01DK082889/DK/NIDDK NIH HHS/United States
- 5T32HL007623/HL/NHLBI NIH HHS/United States
- U01 HL100001/HL/NHLBI NIH HHS/United States
- UO1-HL100001/HL/NHLBI NIH HHS/United States
- K01 DK096013/DK/NIDDK NIH HHS/United States
- 1K01DK096013/DK/NIDDK NIH HHS/United States
- R01 DK082889/DK/NIDDK NIH HHS/United States
- T32 HL007623/HL/NHLBI NIH HHS/United States
- 2T32HL066987/HL/NHLBI NIH HHS/United States
- P50 HG005550/HG/NHGRI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R24 DK092760/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
