

Dnmt3a and Dnmt3b have overlapping and distinct functions in hematopoietic stem cells
- PMID: 25130491
- PMCID: PMC4163922
- DOI: 10.1016/j.stem.2014.06.018
Dnmt3a and Dnmt3b have overlapping and distinct functions in hematopoietic stem cells
Abstract
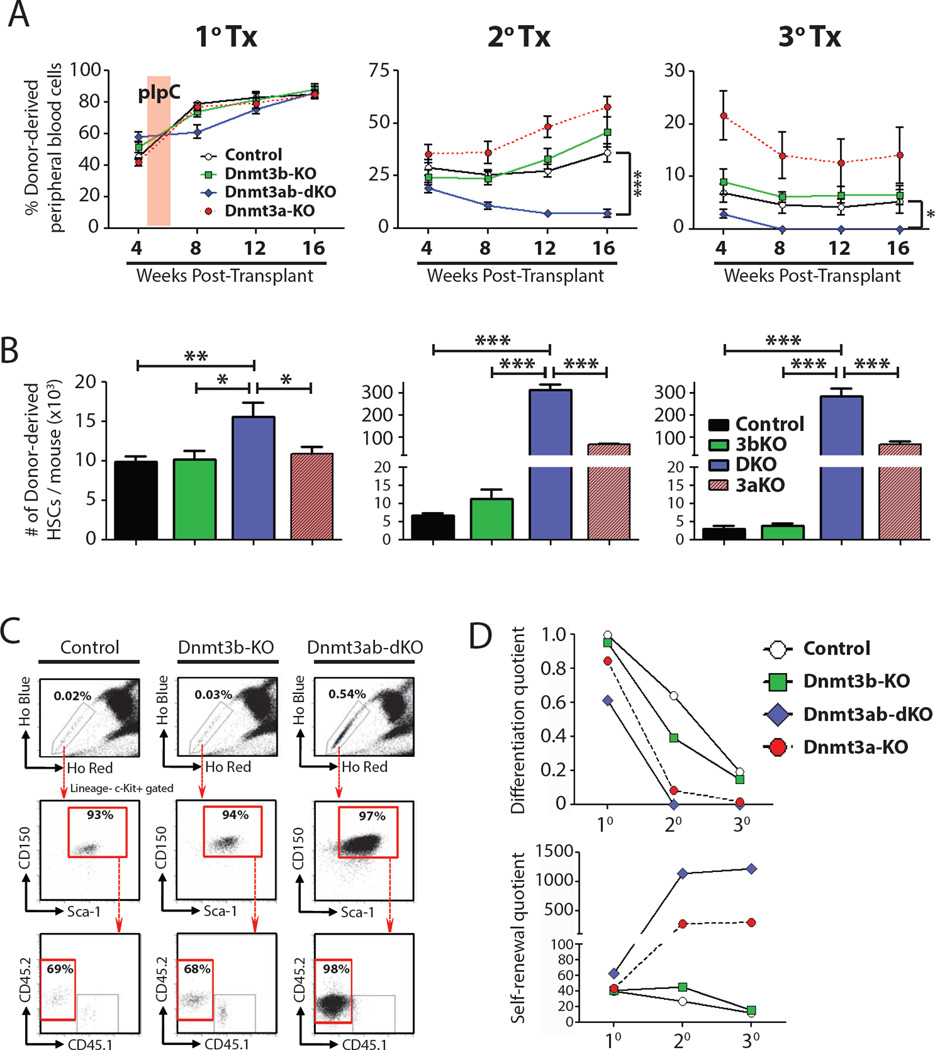
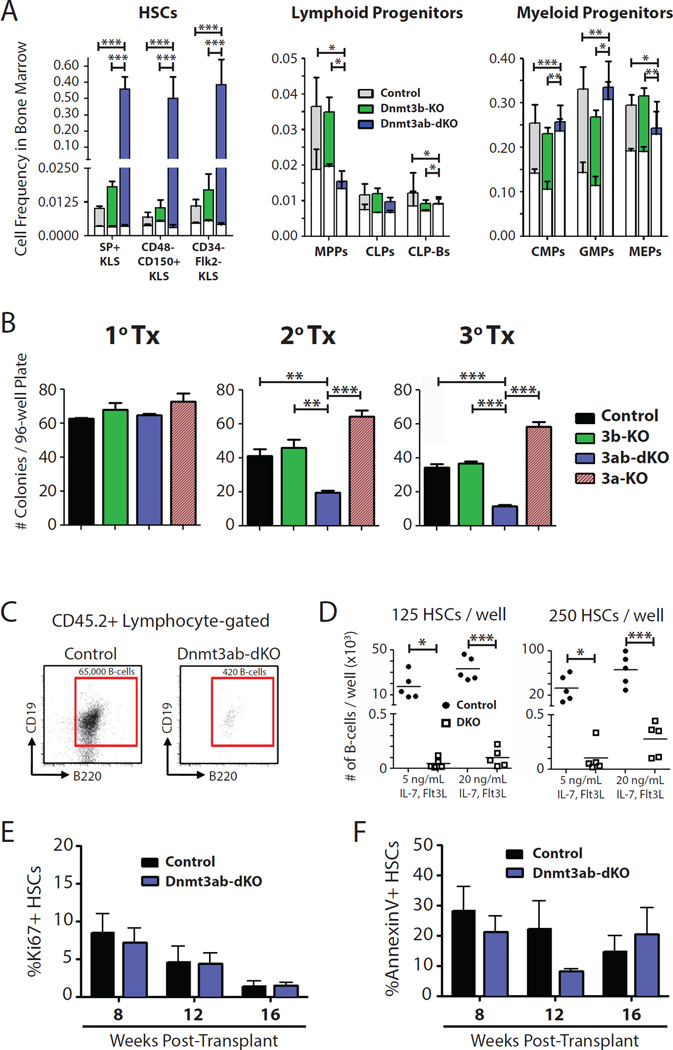
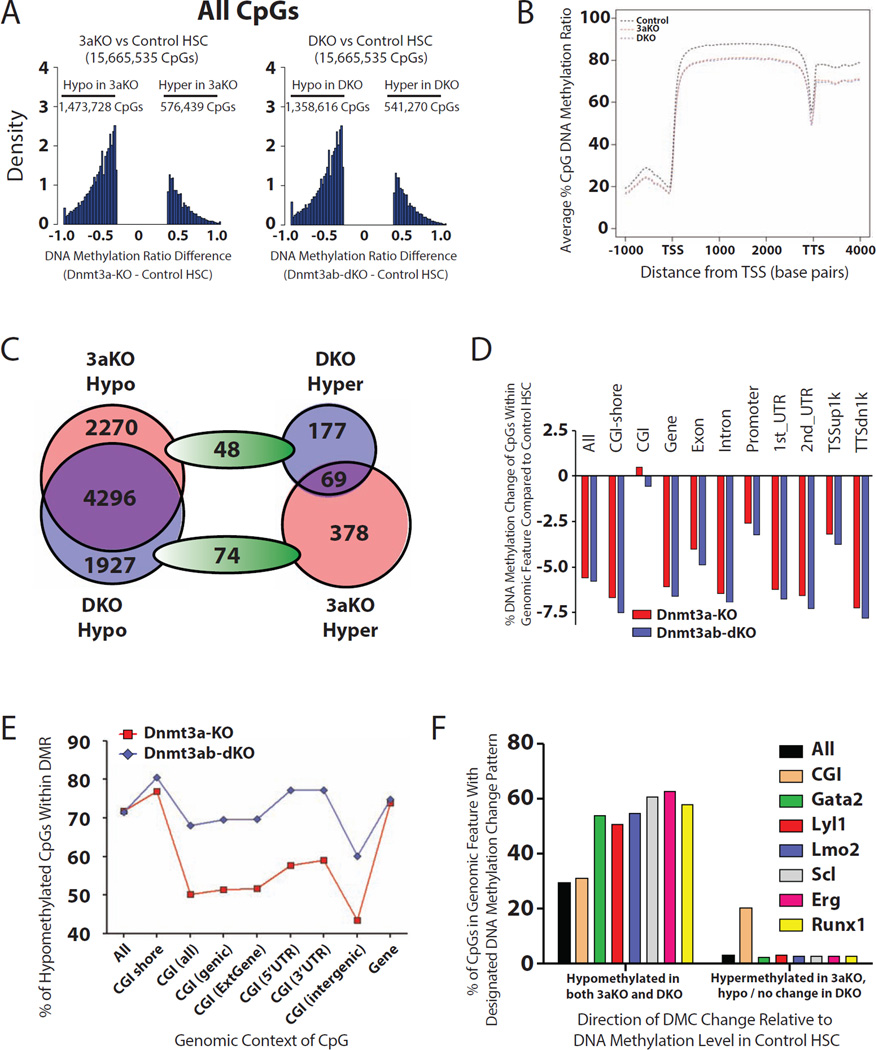
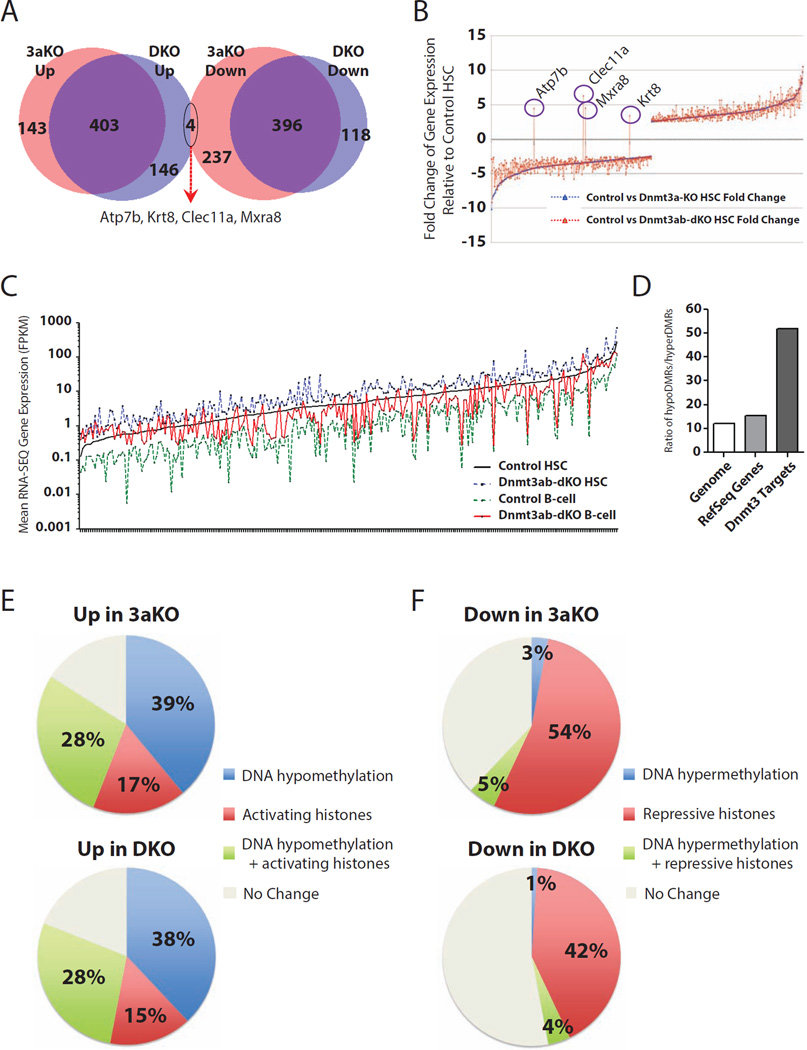
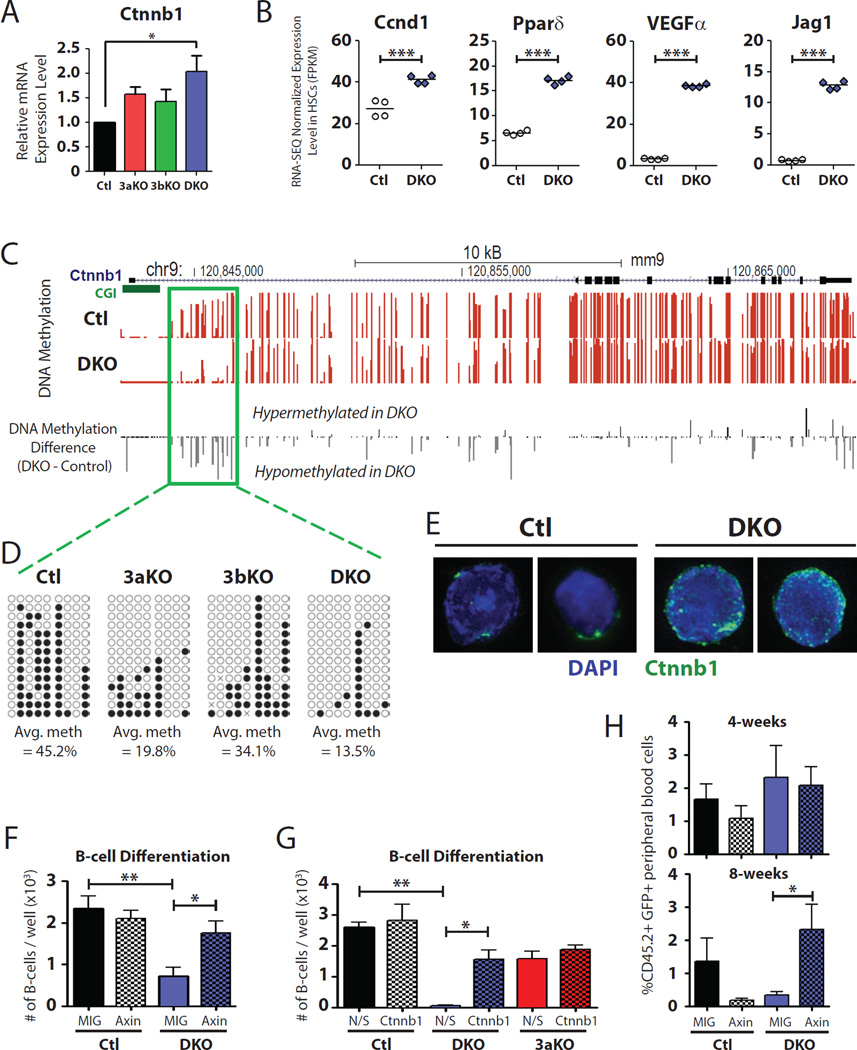
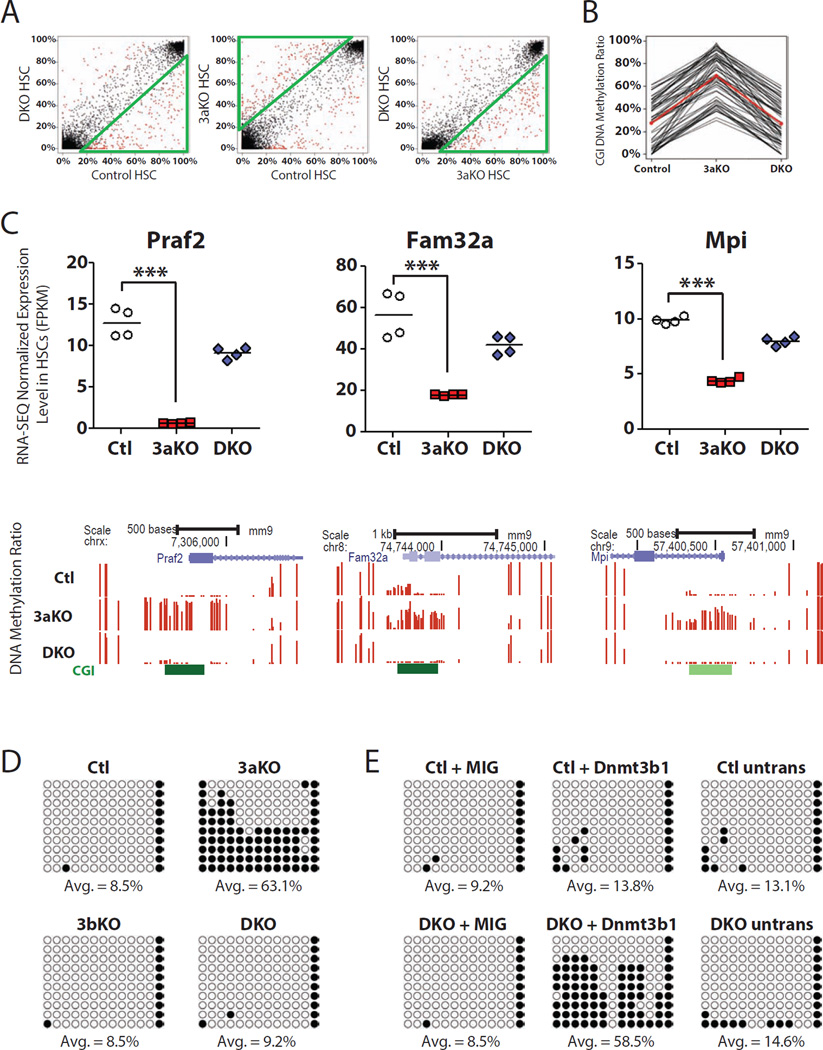
Epigenetic regulation of hematopoietic stem cells (HSCs) ensures lifelong production of blood and bone marrow. Recently, we reported that loss of de novo DNA methyltransferase Dnmt3a results in HSC expansion and impaired differentiation. Here, we report conditional inactivation of Dnmt3b in HSCs either alone or combined with Dnmt3a deletion. Combined loss of Dnmt3a and Dnmt3b was synergistic, resulting in enhanced HSC self-renewal and a more severe block in differentiation than in Dnmt3a-null cells, whereas loss of Dnmt3b resulted in a mild phenotype. Although the predominant Dnmt3b isoform in adult HSCs is catalytically inactive, its residual activity in Dnmt3a-null HSCs can drive some differentiation and generates paradoxical hypermethylation of CpG islands. Dnmt3a/Dnmt3b-null HSCs displayed activated β-catenin signaling, partly accounting for the differentiation block. These data demonstrate distinct roles for Dnmt3b in HSC differentiation and provide insights into complementary de novo methylation patterns governing regulation of HSC fate decisions.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures

References
-
- Beerman I, Bock C, Garrison BS, Smith ZD, Gu H, Meissner A, Rossi DJ. Proliferation-dependent alterations of the DNA methylation landscape underlie hematopoietic stem cell aging. Cell Stem Cell. 2013;12:413–425. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CA125123/CA/NCI NIH HHS/United States
- AI07495/AI/NIAID NIH HHS/United States
- DK092883/DK/NIDDK NIH HHS/United States
- DK084259/DK/NIDDK NIH HHS/United States
- T32 AI007495/AI/NIAID NIH HHS/United States
- P50 CA126752/CA/NCI NIH HHS/United States
- P30 CA125123/CA/NCI NIH HHS/United States
- CA126752/CA/NCI NIH HHS/United States
- RC2 AG036562/AG/NIA NIH HHS/United States
- T32 GM008307/GM/NIGMS NIH HHS/United States
- T32 DK060445/DK/NIDDK NIH HHS/United States
- K99 DK084259/DK/NIDDK NIH HHS/United States
- R56 DK092883/DK/NIDDK NIH HHS/United States
- T32 HL092332/HL/NHLBI NIH HHS/United States
- AG036562/AG/NIA NIH HHS/United States
- R01HG007538/HG/NHGRI NIH HHS/United States
- R01 CA183252/CA/NCI NIH HHS/United States
- R01 HG007538/HG/NHGRI NIH HHS/United States
- R00 DK084259/DK/NIDDK NIH HHS/United States
- R01 DK092883/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
