

Evidence against a stem cell origin of new hepatocytes in a common mouse model of chronic liver injury
- PMID: 25131204
- PMCID: PMC4376310
- DOI: 10.1016/j.celrep.2014.07.003
Evidence against a stem cell origin of new hepatocytes in a common mouse model of chronic liver injury
Erratum in
- Cell Rep. 2014 Sep 11;8(5):1607
Abstract
Hepatocytes provide most liver functions, but they can also proliferate and regenerate the liver after injury. However, under some liver injury conditions, particularly chronic liver injury where hepatocyte proliferation is impaired, liver stem cells (LSCs) are thought to replenish lost hepatocytes. Conflicting results have been reported about the identity of LSCs and their contribution to liver regeneration. To address this uncertainty, we followed candidate LSC populations by genetic fate tracing in adult mice with chronic liver injury due to a choline-deficient, ethionine-supplemented diet. In contrast to previous studies, we failed to detect hepatocytes derived from biliary epithelial cells or mesenchymal liver cells beyond a negligible frequency. In fact, we failed to detect hepatocytes that were not derived from pre-existing hepatocytes. In conclusion, our findings argue against LSCs, or other nonhepatocyte cell types, providing a backup system for hepatocyte regeneration in this common mouse model of chronic liver injury.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
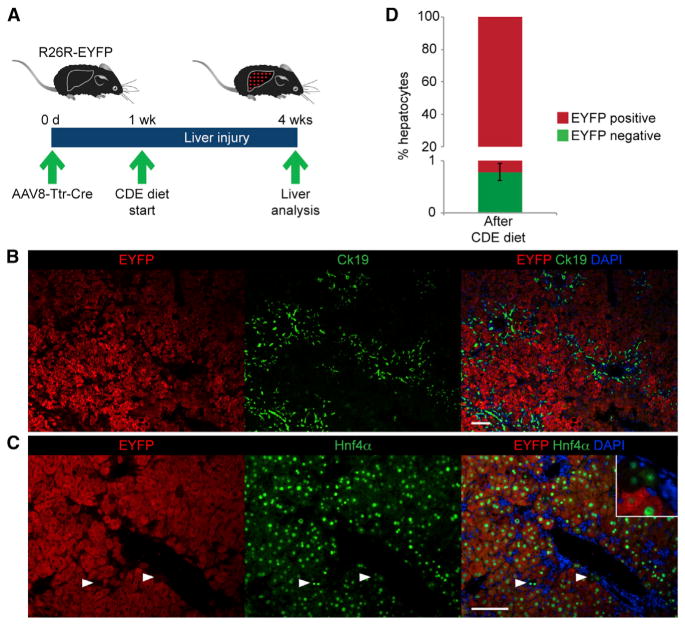
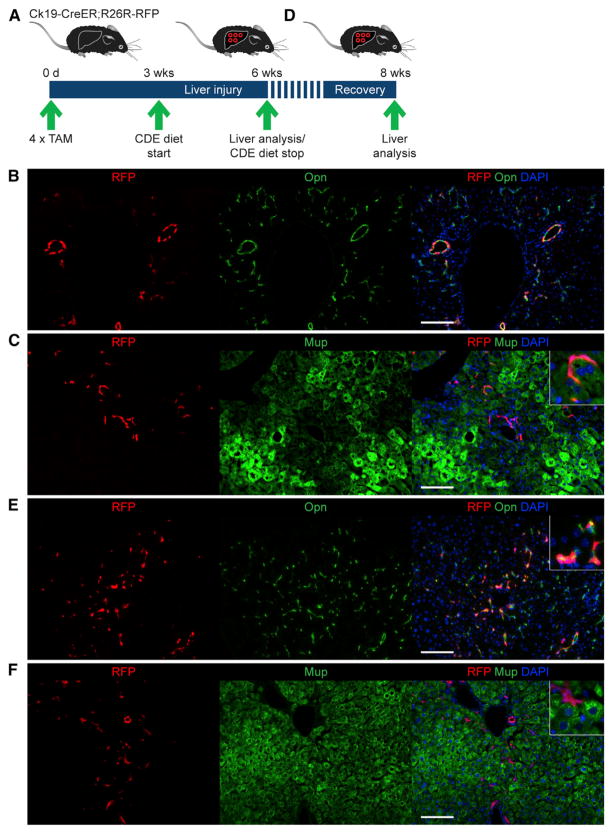
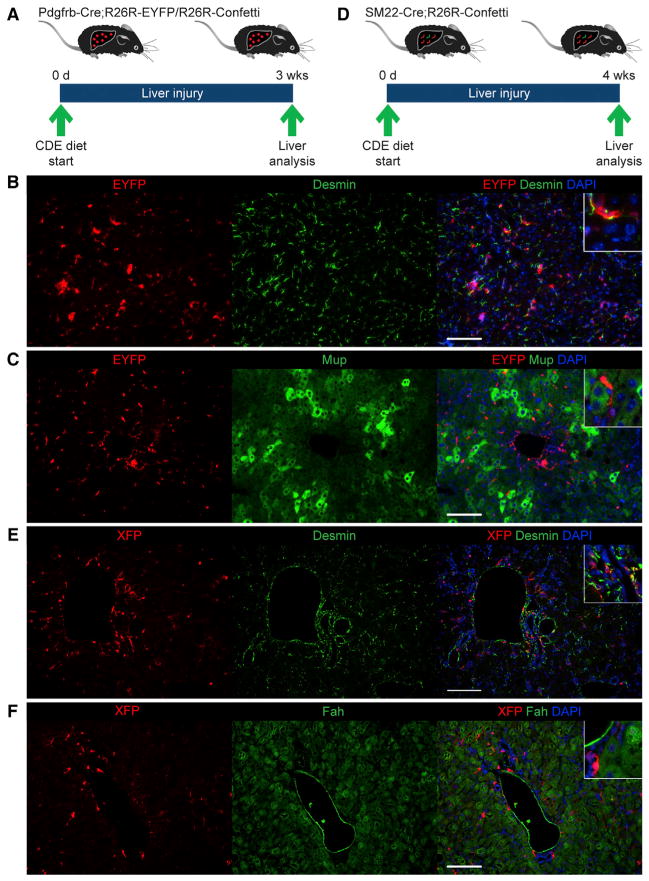
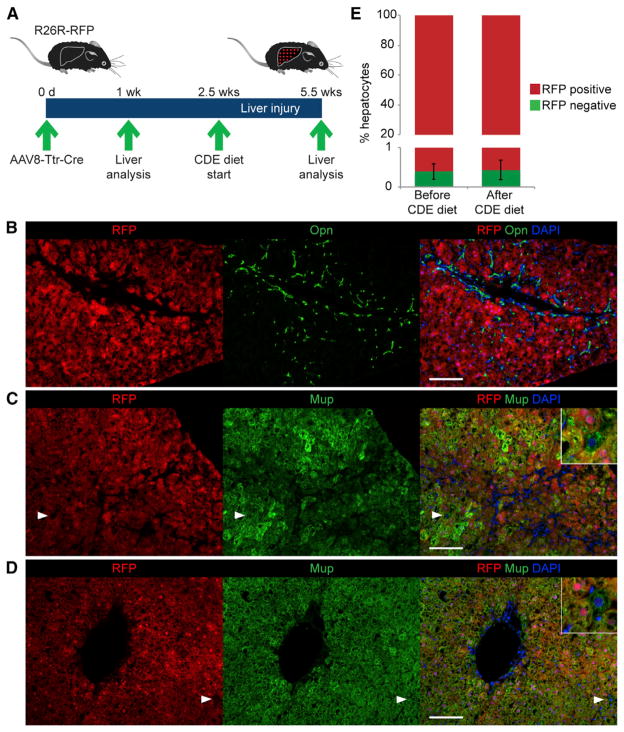
References
-
- Akhurst B, Croager EJ, Farley-Roche CA, Ong JK, Dumble ML, Knight B, Yeoh GC. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology. 2001;34:519–522. - PubMed
-
- Dellavalle A, Sampaolesi M, Tonlorenzi R, Tagliafico E, Sacchetti B, Perani L, Innocenzi A, Galvez BG, Messina G, Morosetti R, et al. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat Cell Biol. 2007;9:255–267. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
