

Stabilization of cartwheel-less centrioles for duplication requires CEP295-mediated centriole-to-centrosome conversion
- PMID: 25131205
- PMCID: PMC4152953
- DOI: 10.1016/j.celrep.2014.07.022
Stabilization of cartwheel-less centrioles for duplication requires CEP295-mediated centriole-to-centrosome conversion
Abstract
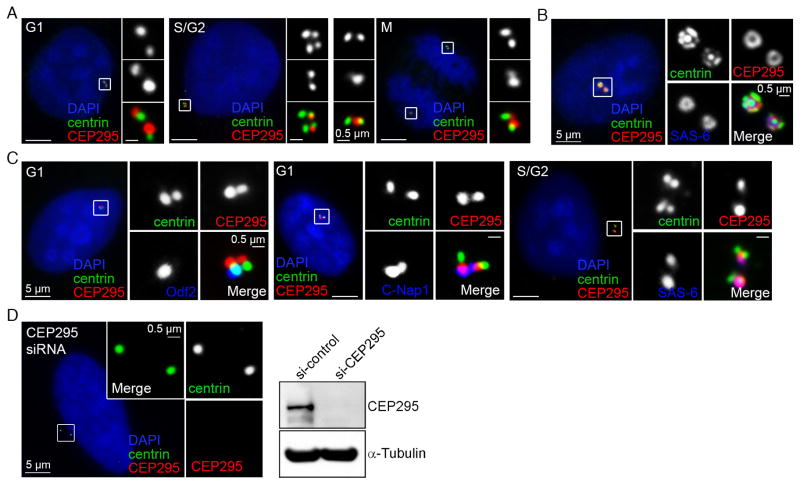
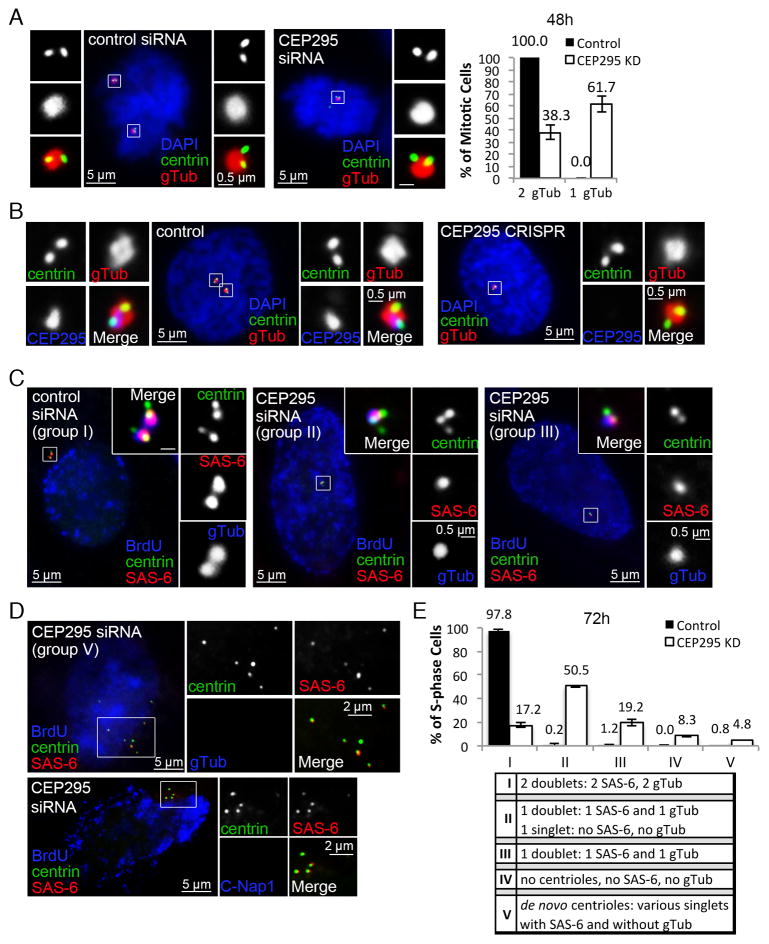
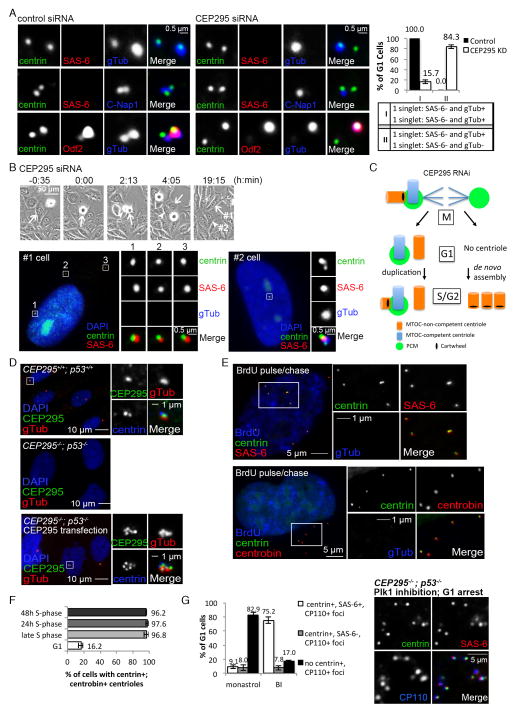
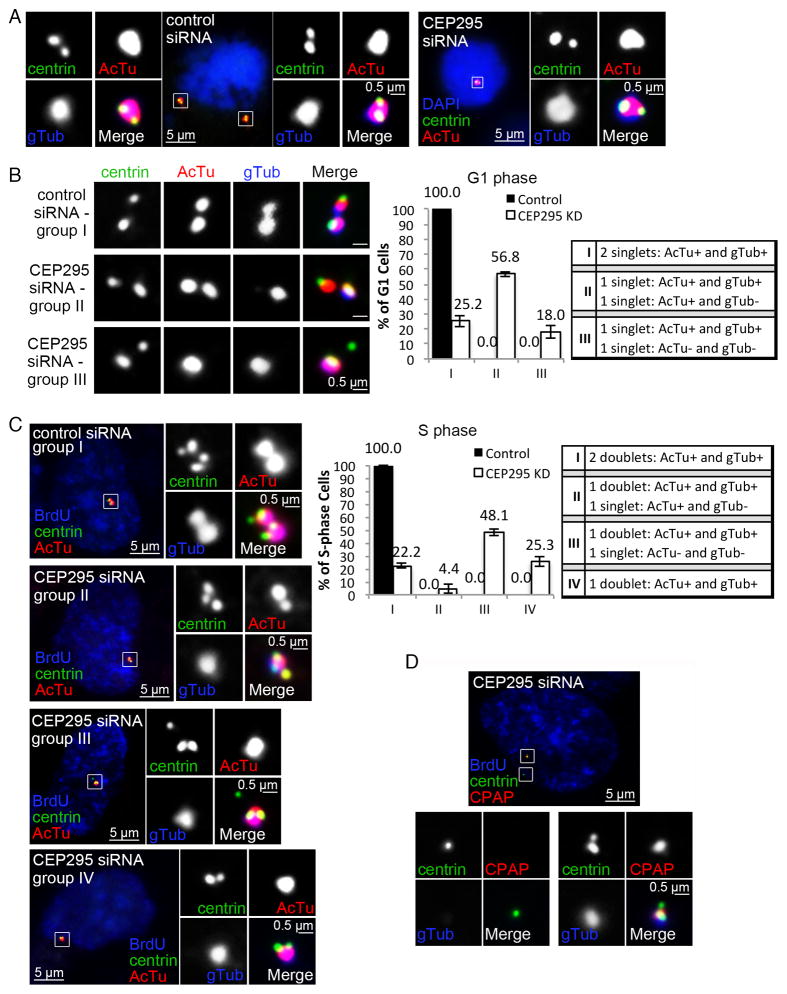
Vertebrate centrioles lose their geometric scaffold, the cartwheel, during mitosis, concurrently with gaining the ability to recruit the pericentriolar material (PCM) and thereby function as the centrosome. Cartwheel removal has recently been implicated in centriole duplication, but whether "cartwheel-less" centrioles are intrinsically stable or must be maintained through other modifications remains unclear. Here, we identify a newborn centriole-enriched protein, KIAA1731/CEP295, specifically mediating centriole-to-centrosome conversion but dispensable for cartwheel removal. In the absence of CEP295, centrioles form in the S/G2 phase and lose their associated cartwheel in mitosis but cannot be converted to centrosomes, uncoupling the two events. Strikingly, centrioles devoid of both the PCM and the cartwheel progressively lose centriolar components, whereas centrioles associating with either the cartwheel or PCM alone can exist stably. Thus, cartwheel removal can have grave repercussions to centriole stability, and centriole-to-centrosome conversion mediated by CEP295 must occur in parallel to maintain cartwheel-less centrioles for duplication.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Arquint C, Nigg EA. STIL microcephaly mutations interfere with APC/C-mediated degradation and cause centriole amplification. Current biology : CB. 2014;24:351–360. - PubMed
-
- Arquint C, Sonnen KF, Stierhof YD, Nigg EA. Cell-cycle-regulated expression of STIL controls centriole number in human cells. Journal of cell science. 2012;125:1342–1352. - PubMed
-
- Fong CS, Kim M, Yang TT, Liao JC, Tsou MF. SAS-6 assembly templated by the lumen of cartwheel-less centrioles precedes centriole duplication. Developmental cell. 2014 http://dx.doi.org/10.1016/j.devcel.2014.1005.1008. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
