

Kcnn4 is a regulator of macrophage multinucleation in bone homeostasis and inflammatory disease
- PMID: 25131209
- PMCID: PMC4471813
- DOI: 10.1016/j.celrep.2014.07.032
Kcnn4 is a regulator of macrophage multinucleation in bone homeostasis and inflammatory disease
Abstract
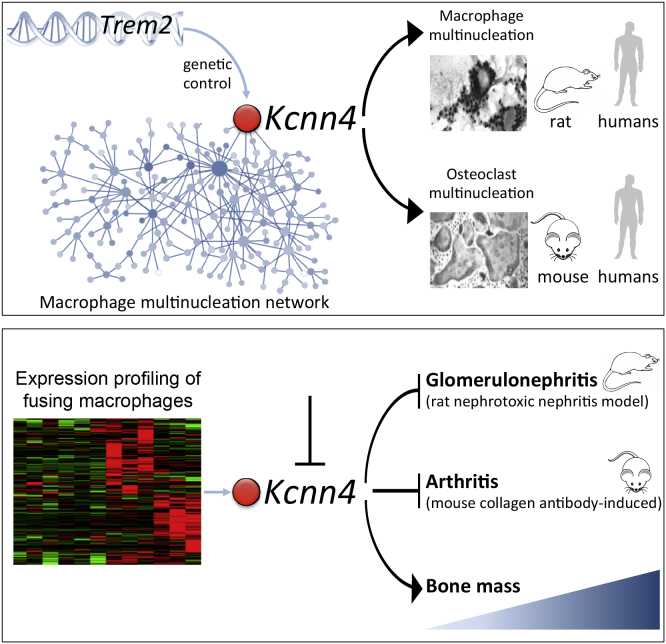
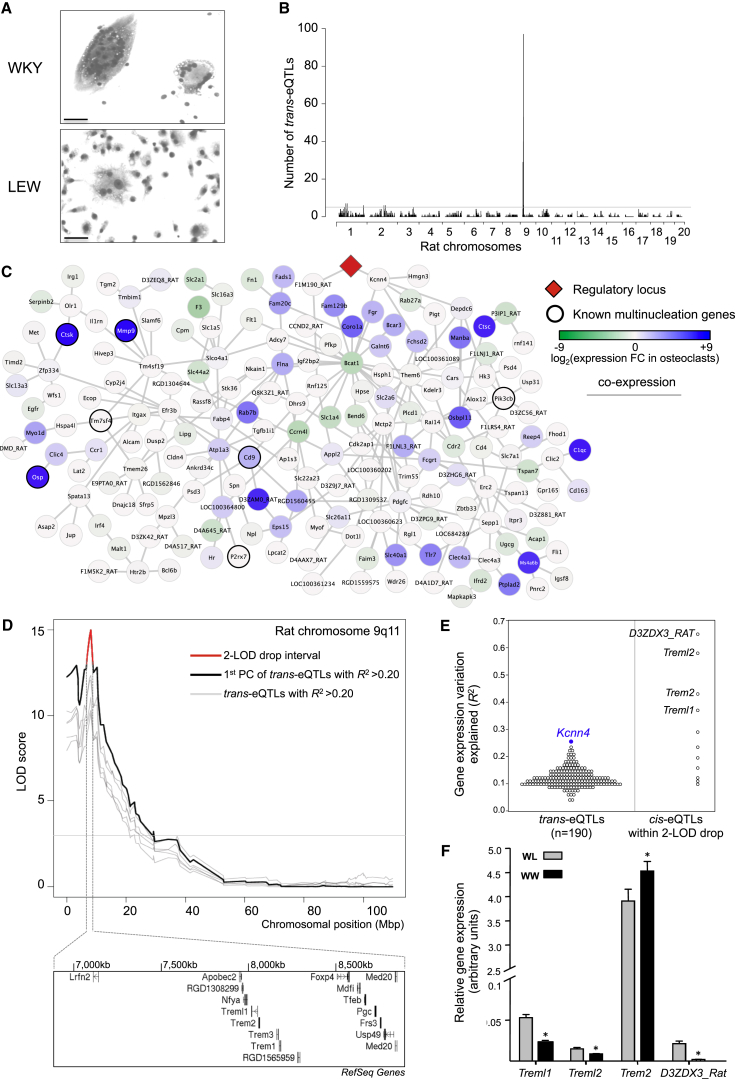
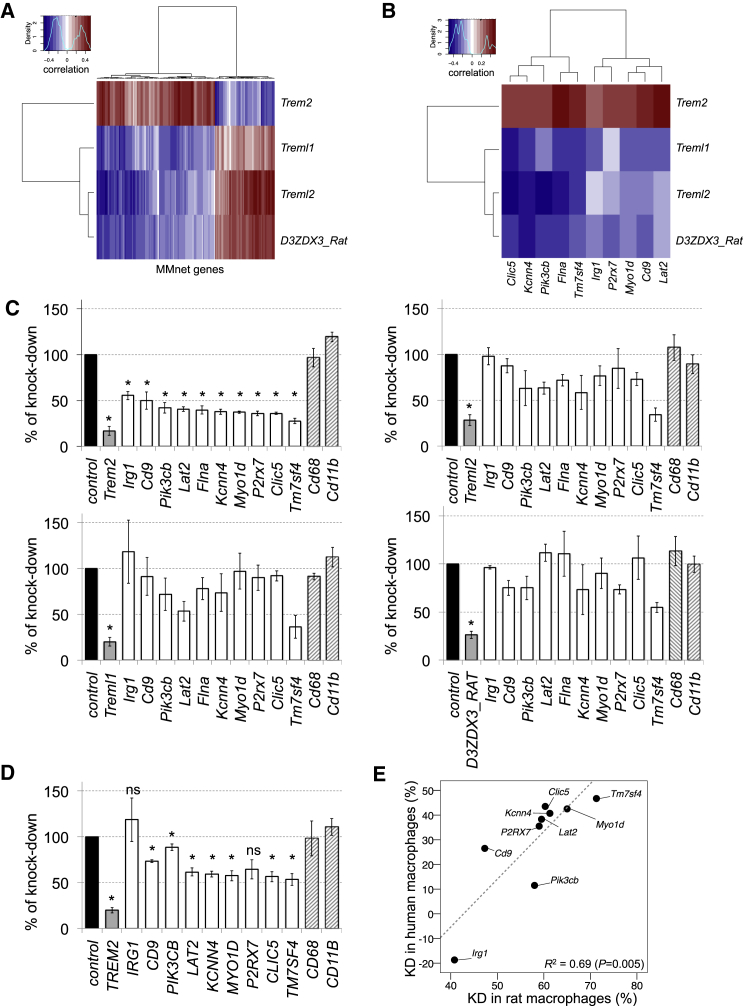
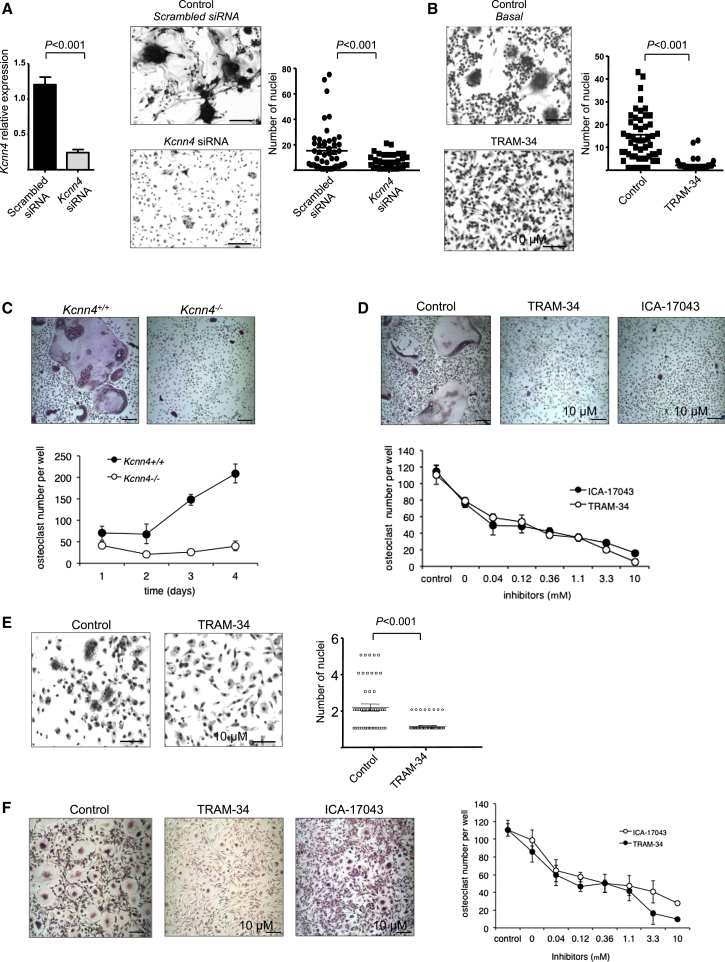
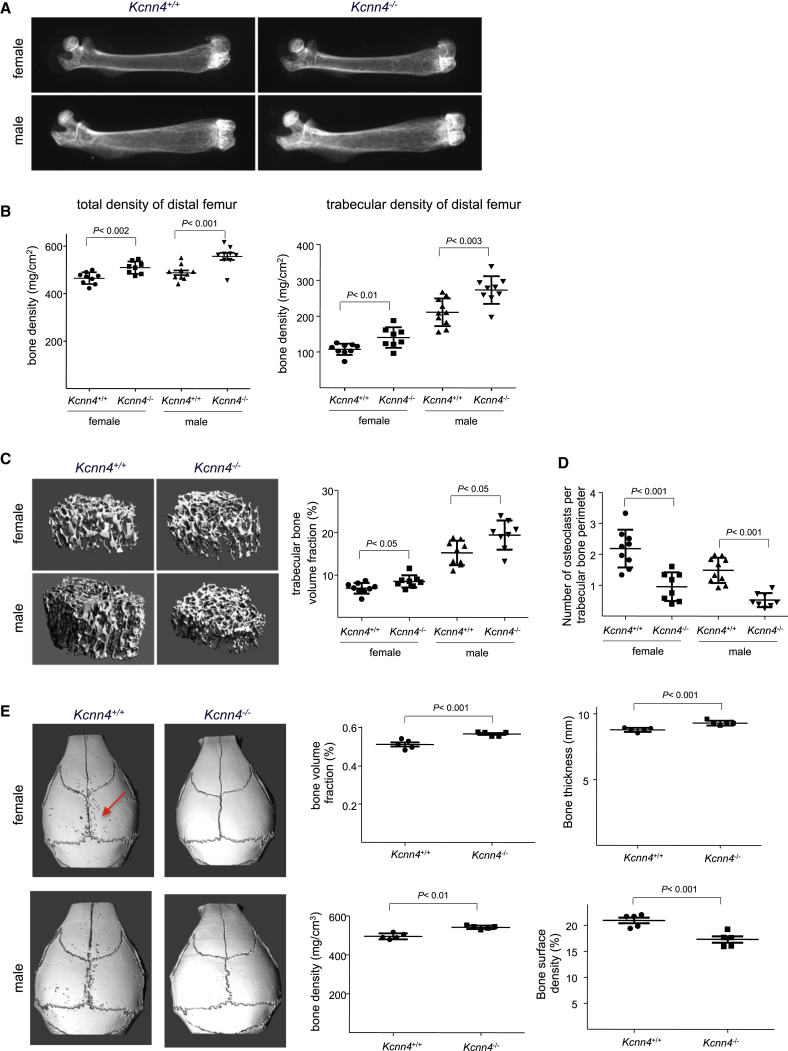
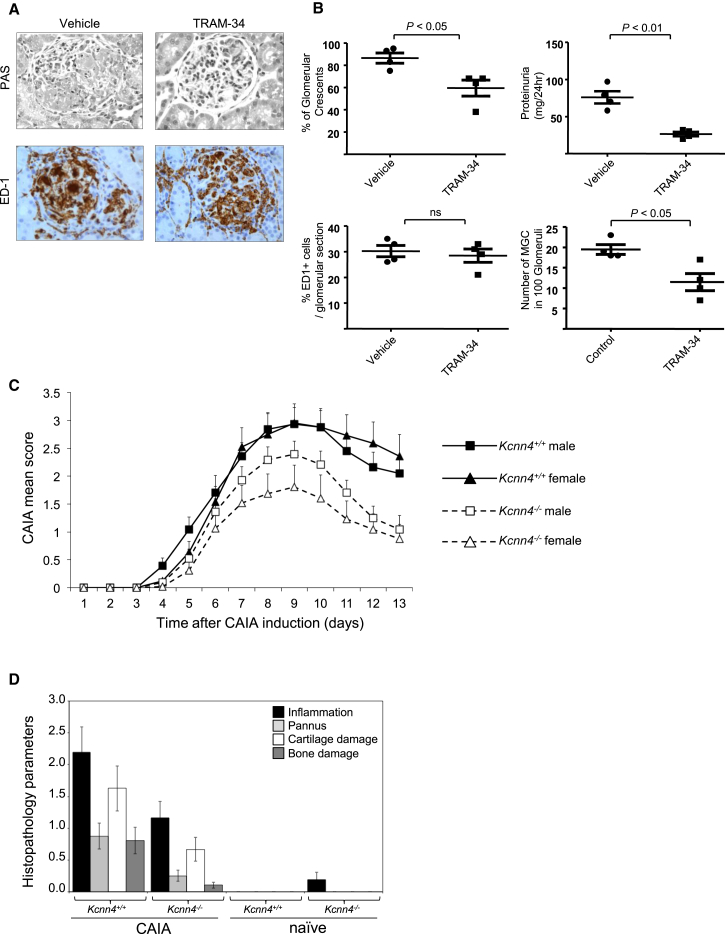
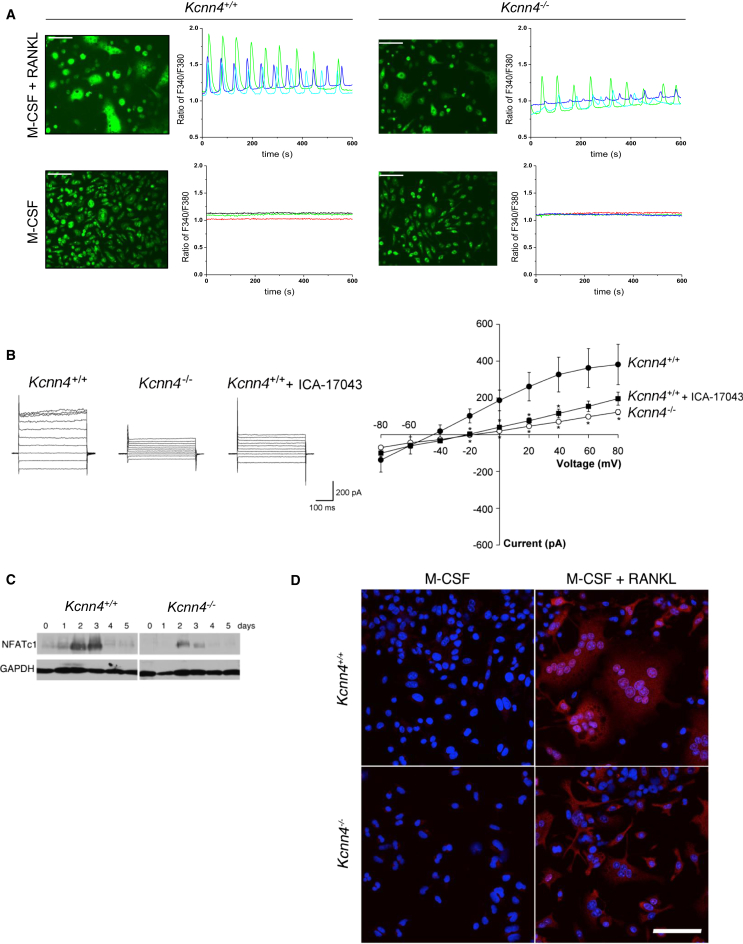
Macrophages can fuse to form osteoclasts in bone or multinucleate giant cells (MGCs) as part of the immune response. We use a systems genetics approach in rat macrophages to unravel their genetic determinants of multinucleation and investigate their role in both bone homeostasis and inflammatory disease. We identify a trans-regulated gene network associated with macrophage multinucleation and Kcnn4 as being the most significantly trans-regulated gene in the network and induced at the onset of fusion. Kcnn4 is required for osteoclast and MGC formation in rodents and humans. Genetic deletion of Kcnn4 reduces macrophage multinucleation through modulation of Ca(2+) signaling, increases bone mass, and improves clinical outcome in arthritis. Pharmacological blockade of Kcnn4 reduces experimental glomerulonephritis. Our data implicate Kcnn4 in macrophage multinucleation, identifying it as a potential therapeutic target for inhibition of bone resorption and chronic inflammation.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Aitman T.J., Dong R., Vyse T.J., Norsworthy P.J., Johnson M.D., Smith J., Mangion J., Roberton-Lowe C., Marshall A.J., Petretto E. Copy number polymorphism in Fcgr3 predisposes to glomerulonephritis in rats and humans. Nature. 2006;439:851–855. - PubMed
-
- Ballica R., Valentijn K., Khachatryan A., Guerder S., Kapadia S., Gundberg C., Gilligan J., Flavell R.A., Vignery A. Targeted expression of calcitonin gene-related peptide to osteoblasts increases bone density in mice. J. Bone Miner. Res. 1999;14:1067–1074. - PubMed
-
- Baron R., Tross R., Vignery A. Evidence of sequential remodeling in rat trabecular bone: morphology, dynamic histomorphometry, and changes during skeletal maturation. Anat. Rec. 1984;208:137–145. - PubMed
-
- Begenisich T., Nakamoto T., Ovitt C.E., Nehrke K., Brugnara C., Alper S.L., Melvin J.E. Physiological roles of the intermediate conductance, Ca2+-activated potassium channel Kcnn4. J. Biol. Chem. 2004;279:47681–47687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
