

Telomerase activation after recruitment in fission yeast
- PMID: 25131669
- PMCID: PMC4159562
- DOI: 10.1016/j.cub.2014.07.035
Telomerase activation after recruitment in fission yeast
Abstract
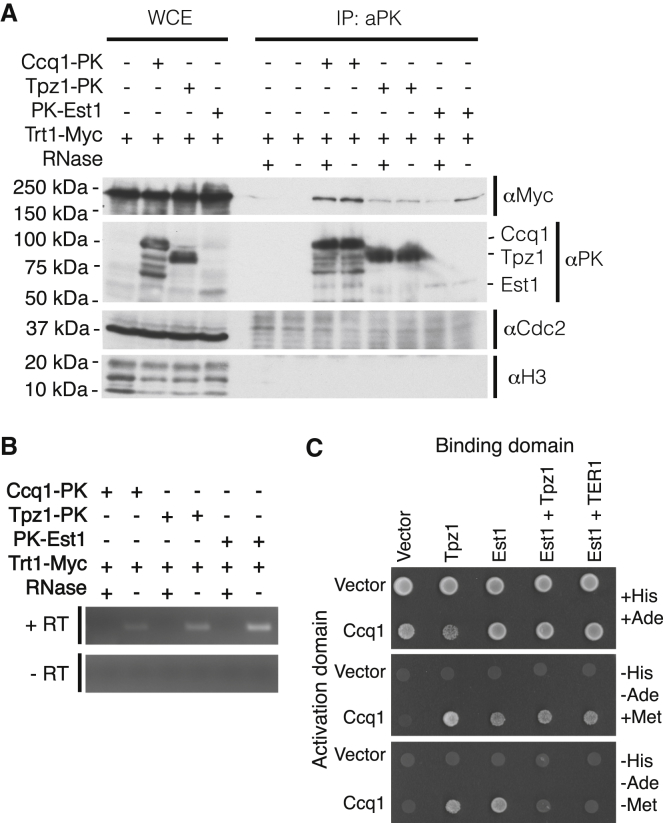
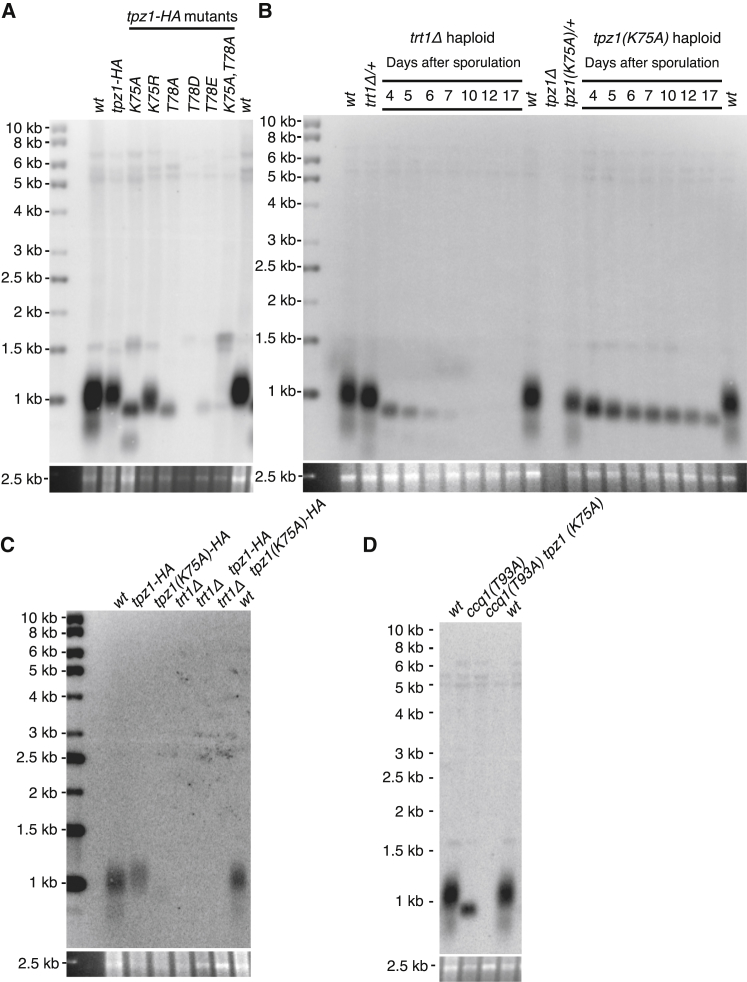
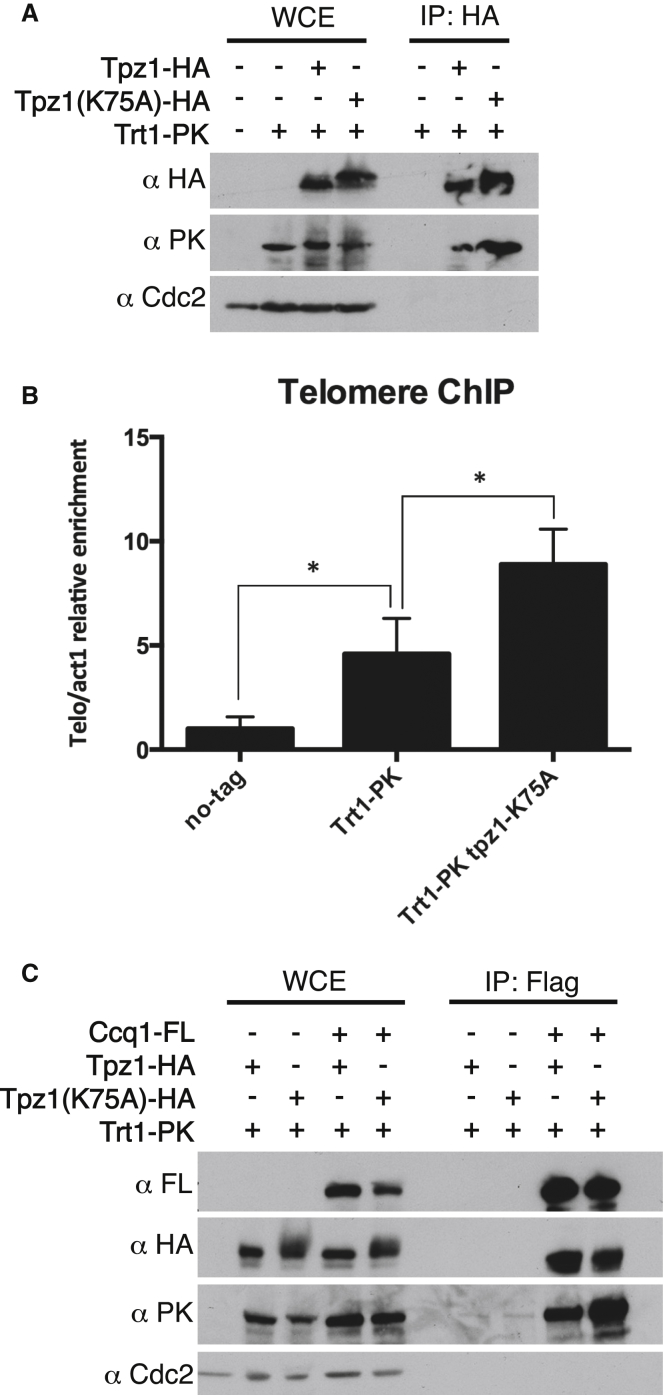
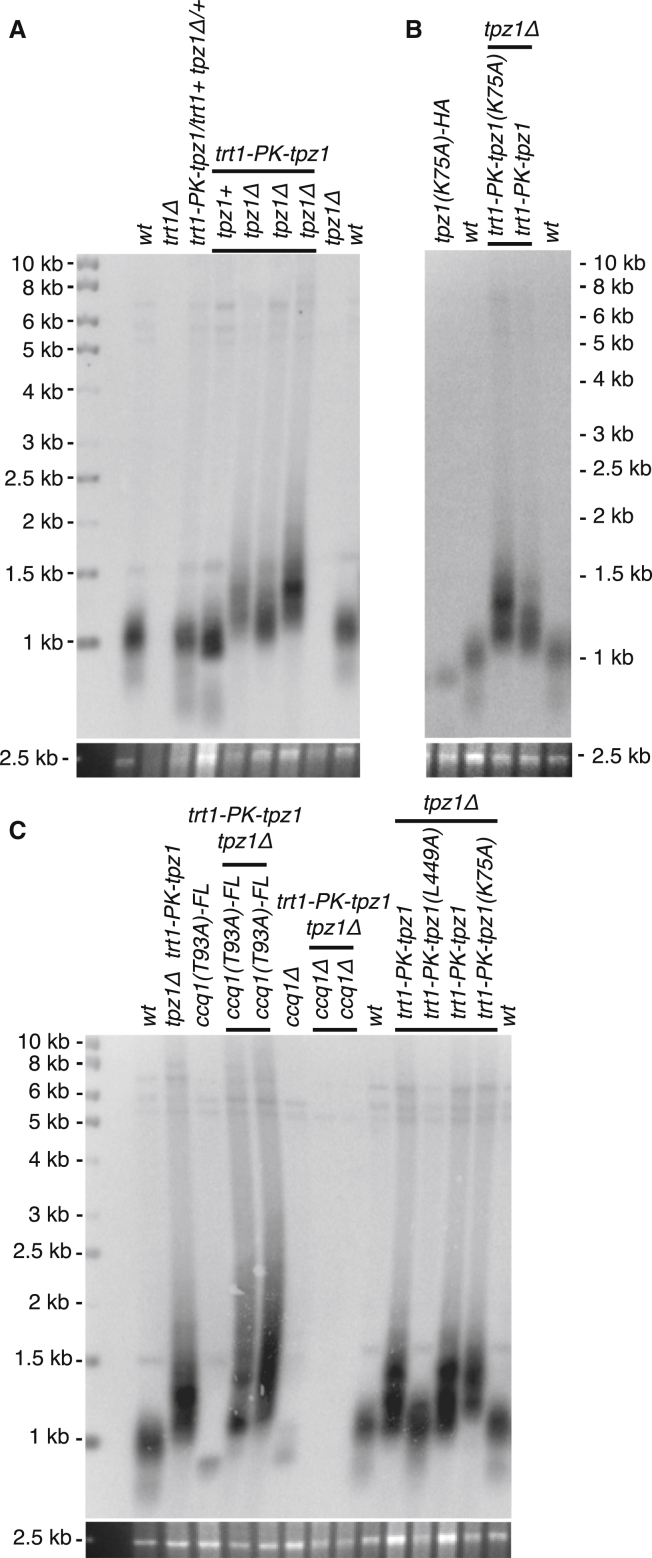
Current models depict that telomerase recruitment equates to activation. Telomeric DNA-binding proteins and the telomerase accessory proteins coordinate the recruitment of telomerase to the ends of chromosomes in a telomere length- and cell-cycle-dependent manner [1-4]. Recent studies have demonstrated that the telomeric protein TPP1 and its binding protein TIN2 are key proteins for both telomerase recruitment and processivity in mammalian cells [5-7]. Although the precise molecular mechanism of telomerase recruitment has not yet been established, targeted point mutations within the oligonucleotide/oligosaccharide-binding (OB)-fold domain of TPP1 have been shown to impair telomerase association and processivity [8-10]. In fission yeast, telomerase is recruited through an interaction between the telomerase subunit Est1 and Ccq1, a component of the Pot1-Tpz1 telomere complex (POT1-TPP1 orthologs) [11-15]. Here, we demonstrate that association of telomerase with telomeres does not engage activity. We describe a mutation of Tpz1 that causes critical telomere shortening despite telomeric accumulation of the telomerase catalytic subunit, Trt1. Furthermore, Est1-directed telomerase association with Ccq1 is transient, and the Est1-Ccq1 interaction does not remain the bridge between telomeres and telomerase. Rather, direct interaction of Trt1 with Tpz1 is critical for telomere elongation. Moreover, Ccq1, which has been well characterized as a telomerase recruiter, is also required for the activation of telomere-associated telomerase. Our findings reveal a layer of telomerase regulation that controls activity after recruitment.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Taggart A.K., Teng S.C., Zakian V.A. Est1p as a cell cycle-regulated activator of telomere-bound telomerase. Science. 2002;297:1023–1026. - PubMed
-
- Teixeira M.T., Arneric M., Sperisen P., Lingner J. Telomere length homeostasis is achieved via a switch between telomerase- extendible and -nonextendible states. Cell. 2004;117:323–335. - PubMed
-
- Wang F., Podell E.R., Zaug A.J., Yang Y., Baciu P., Cech T.R., Lei M. The POT1-TPP1 telomere complex is a telomerase processivity factor. Nature. 2007;445:506–510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
