

HilD induces expression of Salmonella pathogenicity island 2 genes by displacing the global negative regulator H-NS from ssrAB
- PMID: 25135218
- PMCID: PMC4248803
- DOI: 10.1128/JB.01799-14
HilD induces expression of Salmonella pathogenicity island 2 genes by displacing the global negative regulator H-NS from ssrAB
Abstract
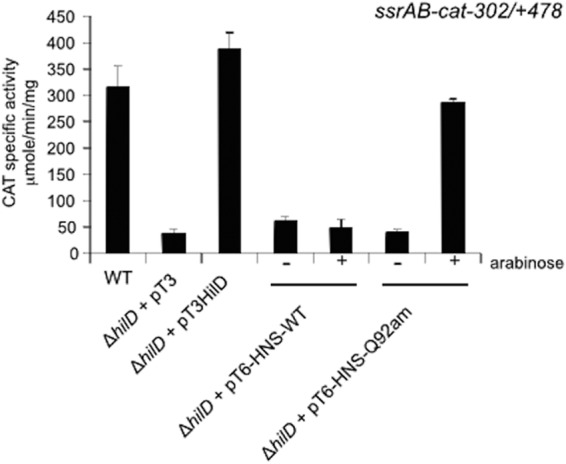
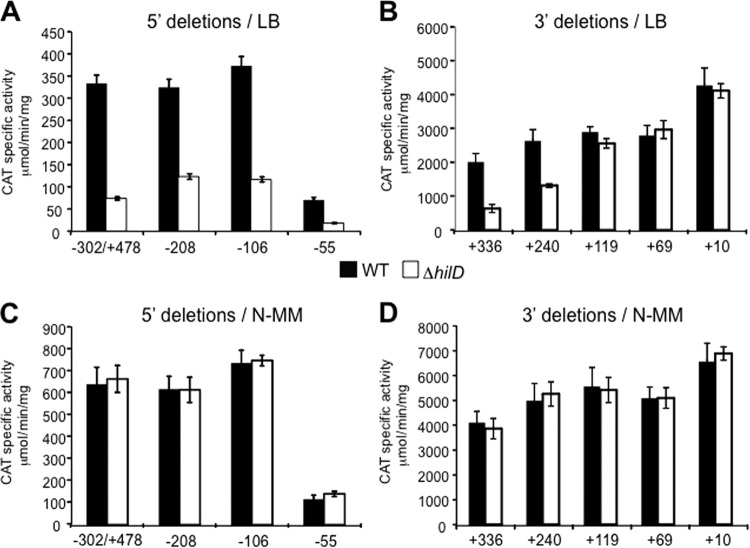
Salmonella pathogenicity islands 1 and 2 (SPI-1 and SPI-2) have essential roles in the pathogenesis of Salmonella enterica. Previously, we reported transcriptional cross talk between SPI-1 and SPI-2 when the SPI-1 regulator HilD induces expression of the SsrA/B two-component system, the central positive regulator of SPI-2, during the growth of Salmonella to late stationary phase in LB rich medium. Here, we further define the mechanism of the HilD-mediated expression of ssrAB. Expression analysis of cat transcriptional fusions containing different regions of ssrAB revealed the presence of negative regulatory sequences located downstream of the ssrAB promoter. In the absence of these negative cis elements, ssrAB was expressed in a HilD-independent manner and was no longer repressed by the global regulator H-NS. Consistently, when the activity of H-NS was inactivated, the expression of ssrAB also became independent of HilD. Furthermore, electrophoretic mobility shift assays showed that both HilD and H-NS bind to the ssrAB region containing the repressing sequences. Moreover, HilD was able to displace H-NS bound to this region, whereas H-NS did not displace HilD. Our results support a model indicating that HilD displaces H-NS from a region downstream of the promoter of ssrAB by binding to sites overlapping or close to those sites bound by H-NS, which leads to the expression of ssrAB. Although the role of HilD as an antagonist of H-NS has been reported before for other genes, this is the first study showing that HilD is able to effectively displace H-NS from the promoter of one of its target genes.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
