

Oxidoreductase activity is necessary for N-glycosylation of cysteine-proximal acceptor sites in glycoproteins
- PMID: 25135935
- PMCID: PMC4137057
- DOI: 10.1083/jcb.201404083
Oxidoreductase activity is necessary for N-glycosylation of cysteine-proximal acceptor sites in glycoproteins
Abstract
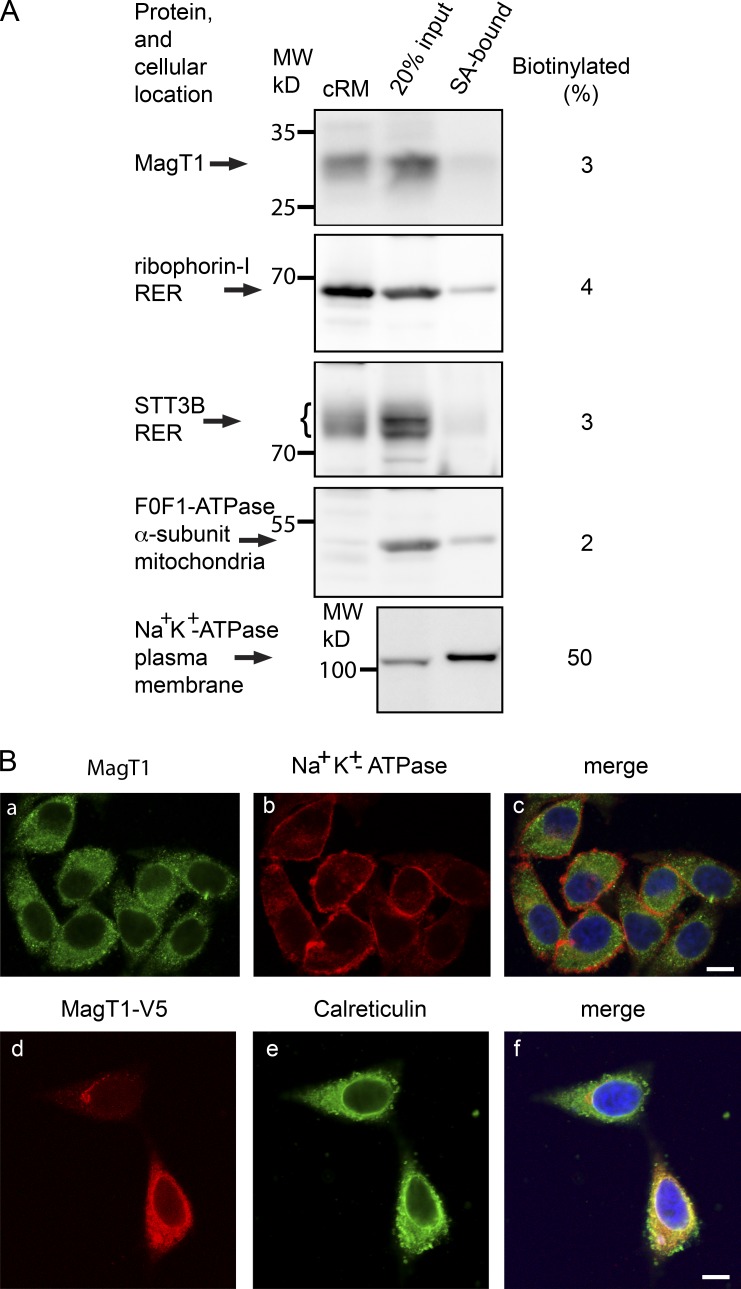
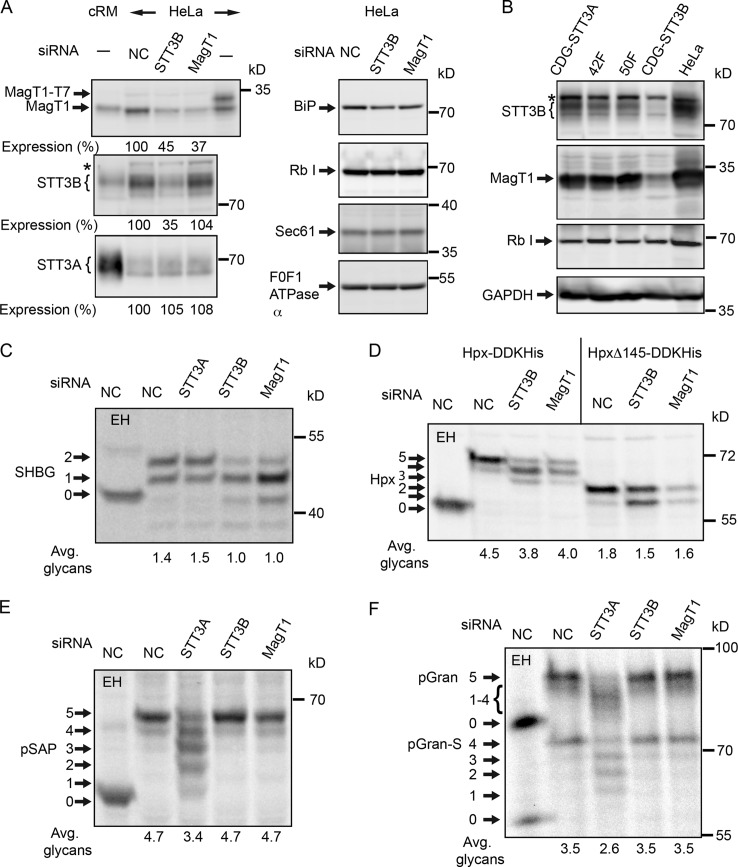
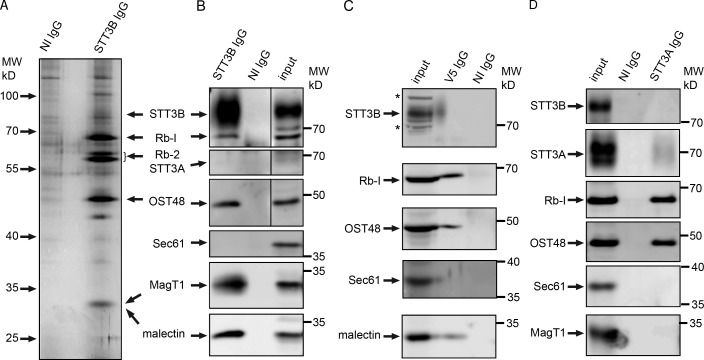
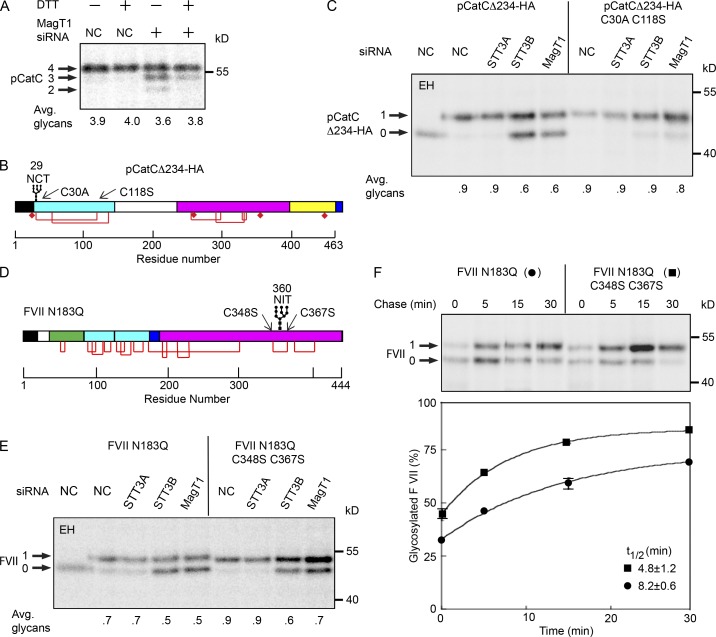
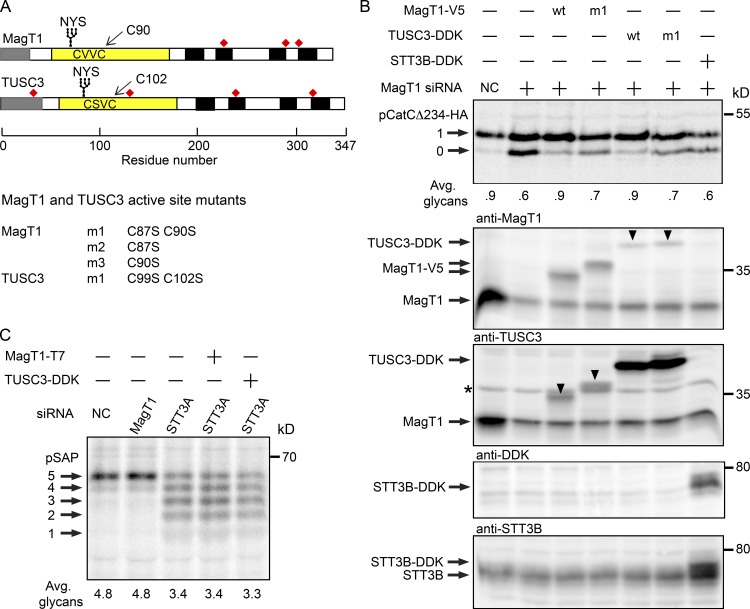
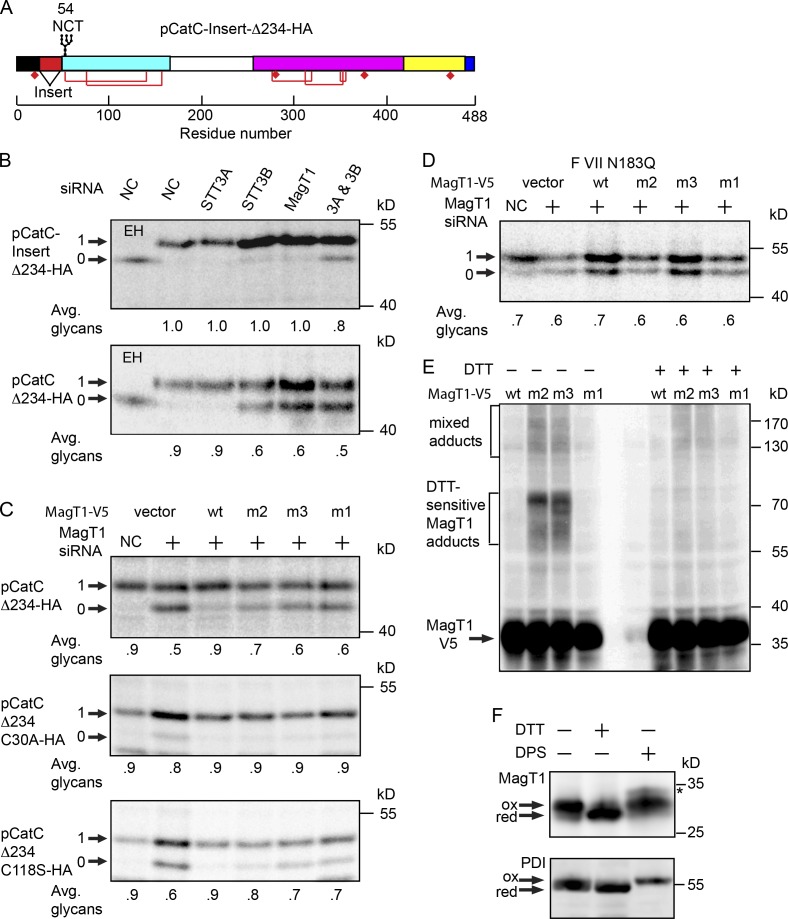
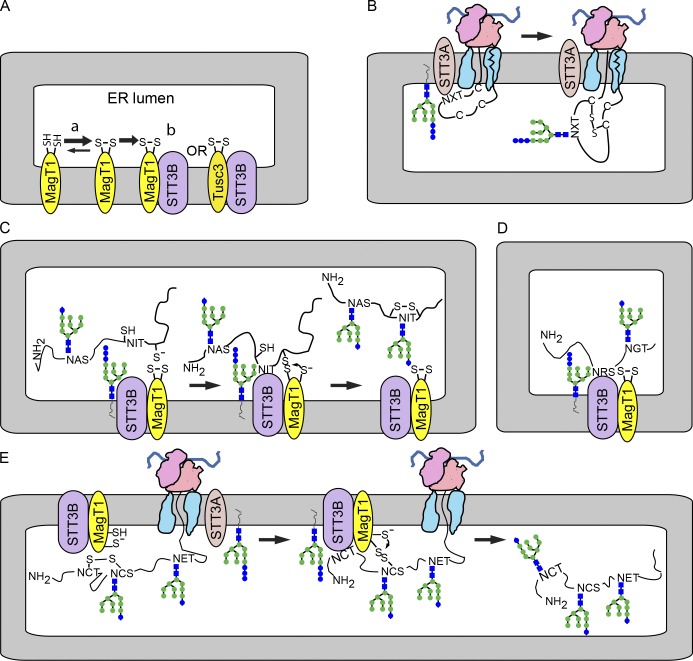
Stabilization of protein tertiary structure by disulfides can interfere with glycosylation of acceptor sites (NXT/S) in nascent polypeptides. Here, we show that MagT1, an ER-localized thioredoxin homologue, is a subunit of the STT3B isoform of the oligosaccharyltransferase (OST). The lumenally oriented active site CVVC motif in MagT1 is required for glycosylation of STT3B-dependent acceptor sites including those that are closely bracketed by disulfides or contain cysteine as the internal residue (NCT/S). The MagT1- and STT3B-dependent glycosylation of cysteine-proximal acceptor sites can be reduced by eliminating cysteine residues. The predominant form of MagT1 in vivo is oxidized, which is consistent with transient formation of mixed disulfides between MagT1 and a glycoprotein substrate to facilitate access of STT3B to unmodified acceptor sites. Cotranslational N-glycosylation by the STT3A isoform of the OST, which lacks MagT1, allows efficient modification of acceptor sites in cysteine-rich protein domains before disulfide bond formation. Thus, mammalian cells use two mechanisms to achieve N-glycosylation of cysteine proximal acceptor sites.
© 2014 Cherepanova et al.
Figures
References
-
- Bocchinfuso, W.P., Ma K.L., Lee W.M., Warmels-Rodenhiser S., and Hammond G.L.. 1992. Selective removal of glycosylation sites from sex hormone-binding globulin by site-directed mutagenesis. Endocrinology. 131:2331–2336 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
