

Extracellular-vesicle type of volume transmission and tunnelling-nanotube type of wiring transmission add a new dimension to brain neuro-glial networks
- PMID: 25135966
- PMCID: PMC4142026
- DOI: 10.1098/rstb.2013.0505
Extracellular-vesicle type of volume transmission and tunnelling-nanotube type of wiring transmission add a new dimension to brain neuro-glial networks
Abstract
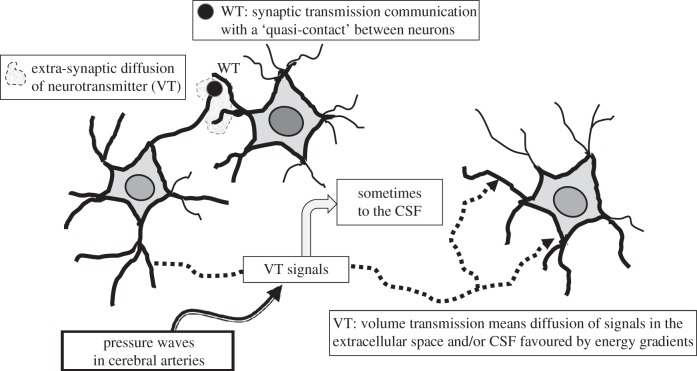
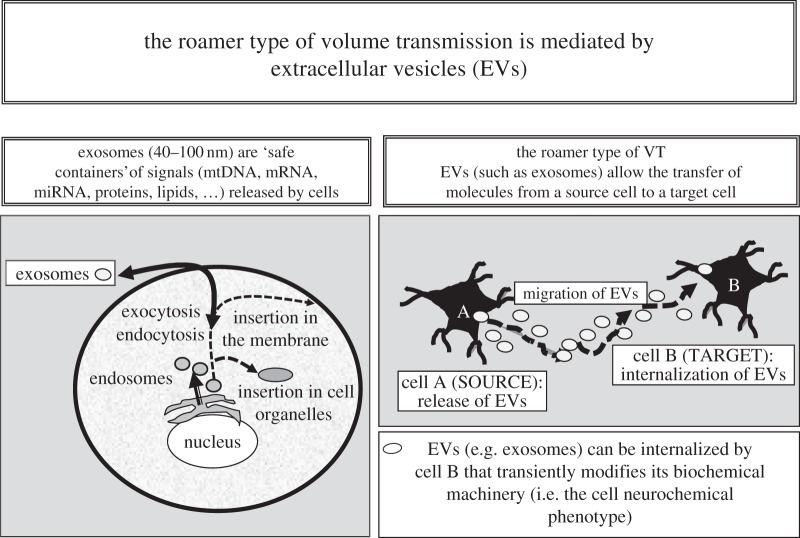
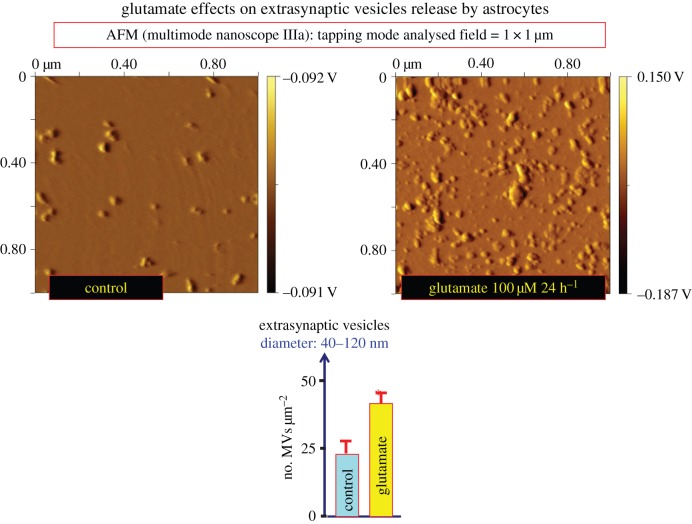
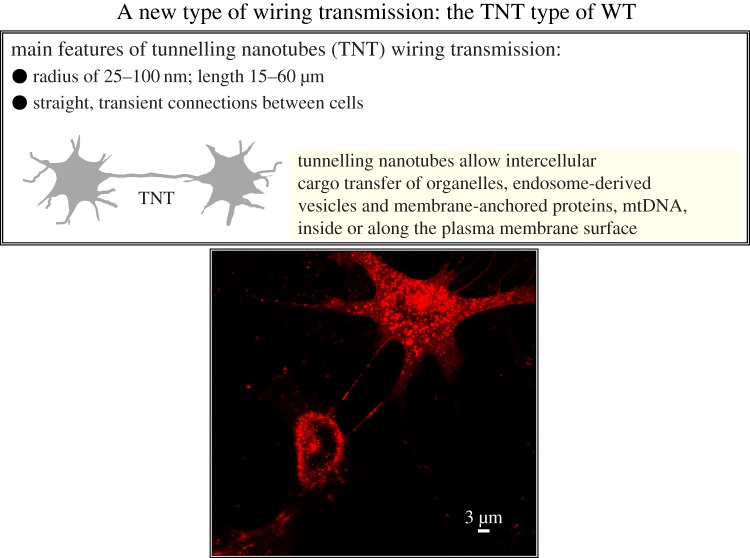
Two major types of intercellular communication are found in the central nervous system (CNS), namely wiring transmission (WT; point-to-point communication via private channels, e.g. synaptic transmission) and volume transmission (VT; communication in the extracellular fluid and in the cerebrospinal fluid). Volume and synaptic transmission become integrated because their chemical signals activate different types of interacting receptors in heteroreceptor complexes located synaptically and extrasynaptically in the plasma membrane. In VT, we focus on the role of the extracellular-vesicle type of VT, and in WT, on the potential role of the tunnelling-nanotube (TNT) type of WT. The so-called exosomes appear to be the major vesicular carrier for intercellular communication but the larger microvesicles also participate. Extracellular vesicles are released from cultured cortical neurons and different types of glial cells and modulate the signalling of the neuronal-glial networks of the CNS. This type of VT has pathological relevance, and epigenetic mechanisms may participate in the modulation of extracellular-vesicle-mediated VT. Gerdes and co-workers proposed the existence of a novel type of WT based on TNTs, which are straight transcellular channels leading to the formation in vitro of syncytial cellular networks found also in neuronal and glial cultures.
Keywords: exosomes; extracellular vesicles; microvesicles; tunnelling nanotubes; volume transmission; wiring transmission.
© 2014 The Author(s) Published by the Royal Society. All rights reserved.
Figures
References
-
- Agnati LF, Fuxe K, Zoli M, Ozini I, Toffano G, Ferraguti F. 1986. A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol. Scand. 128, 201–207. (10.1111/j.1748-1716.1986.tb07967.x) - DOI - PubMed
-
- Fuxe K, Agnati LF, Zoli M, Cintra A, Harfstrand A, von Euler G, Grimaldi R, Kalia M, Eneroth P. 1988. The opioid peptide systems: their organization and role in volume transmission and neuroendocrine regulation. In Regulatory roles of opioid peptides (eds Illes P, Farsang C.), pp. 33–68. Weinheim, Germany: VCH.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
