

Epigenetic setting and reprogramming for neural cell fate determination and differentiation
- PMID: 25135972
- PMCID: PMC4142032
- DOI: 10.1098/rstb.2013.0511
Epigenetic setting and reprogramming for neural cell fate determination and differentiation
Abstract
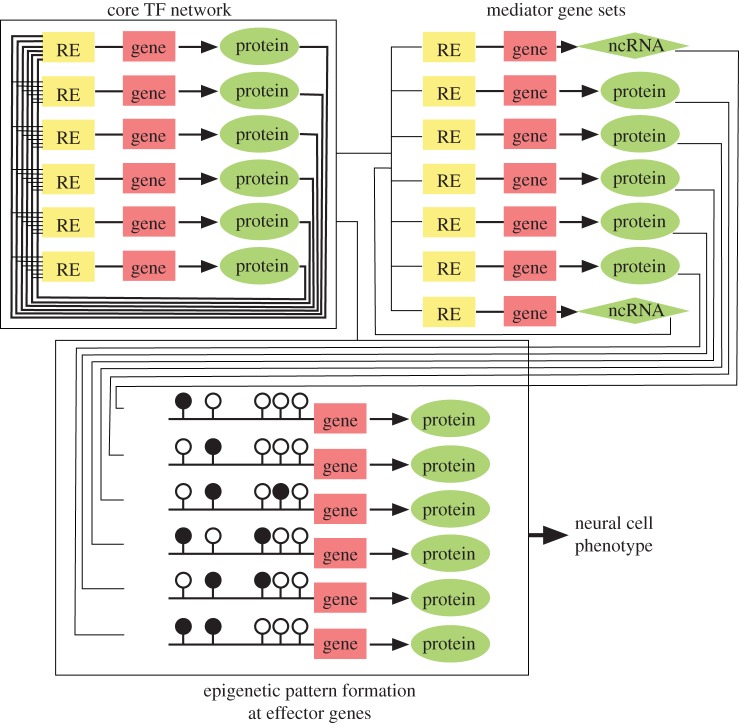
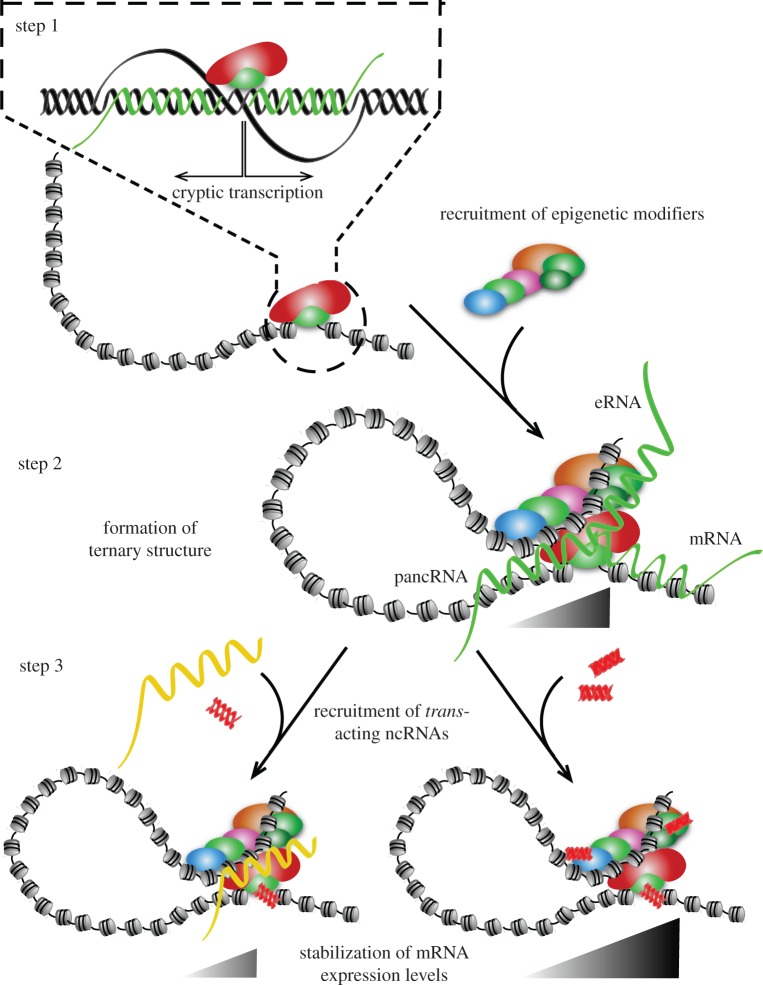
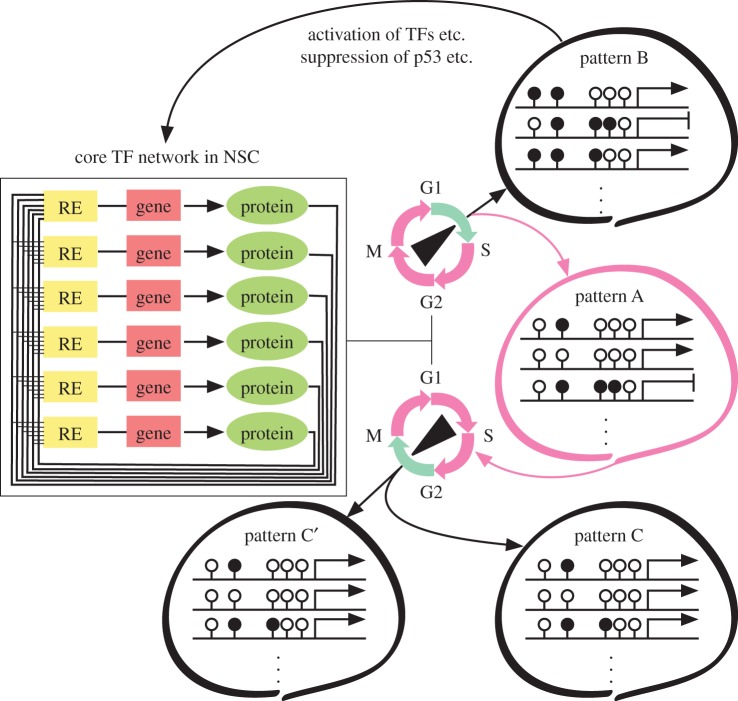
In the mammalian brain, epigenetic mechanisms are clearly involved in the regulation of self-renewal of neural stem cells and the derivation of their descendants, i.e. neurons, astrocytes and oligodendrocytes, according to the developmental timing and the microenvironment, the 'niche'. Interestingly, local epigenetic changes occur, concomitantly with genome-wide level changes, at a set of gene promoter regions for either down- or upregulation of the gene. In addition, intergenic regions also sensitize the availability of epigenetic modifiers, which affects gene expression through a relatively long-range chromatinic interaction with the transcription regulatory machineries including non-coding RNA (ncRNA) such as promoter-associated ncRNA and enhancer ncRNA. We show that such an epigenetic landscape in a neural cell is statically but flexibly formed together with a variable combination of generally and locally acting nuclear molecules including master transcription factors and cell-cycle regulators. We also discuss the possibility that revealing the epigenetic regulation by the local DNA-RNA-protein assemblies would promote methodological innovations, e.g. neural cell reprogramming, engineering and transplantation, to manipulate neuronal and glial cell fates for the purpose of medical use of these cells.
Keywords: DNA methylation; REST; histone acetylation; histone methylation; non-coding RNA; polycomb repressive complex 2.
© 2014 The Author(s) Published by the Royal Society. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources