

Transcriptome analysis of the oil-rich tea plant, Camellia oleifera, reveals candidate genes related to lipid metabolism
- PMID: 25136805
- PMCID: PMC4138098
- DOI: 10.1371/journal.pone.0104150
Transcriptome analysis of the oil-rich tea plant, Camellia oleifera, reveals candidate genes related to lipid metabolism
Abstract
Background: Rapidly driven by the need for developing sustainable sources of nutritionally important fatty acids and the rising concerns about environmental impacts after using fossil oil, oil-plants have received increasing awareness nowadays. As an important oil-rich plant in China, Camellia oleifera has played a vital role in providing nutritional applications, biofuel productions and chemical feedstocks. However, the lack of C. oleifera genome sequences and little genetic information have largely hampered the urgent needs for efficient utilization of the abundant germplasms towards modern breeding efforts of this woody oil-plant.
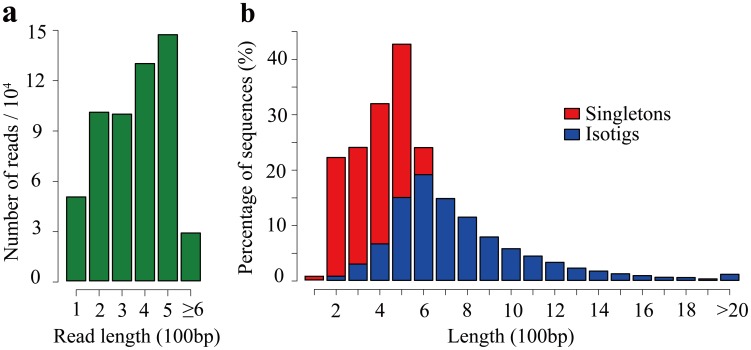
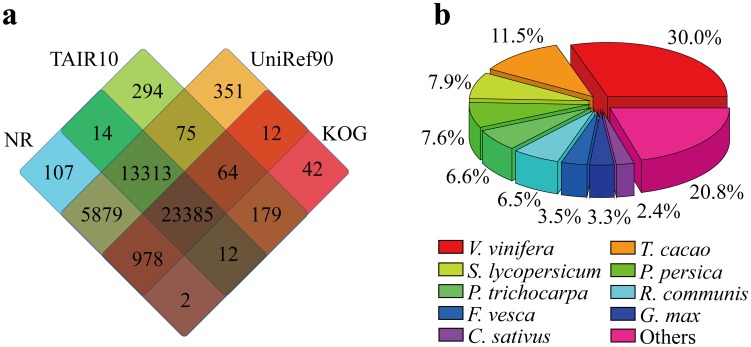
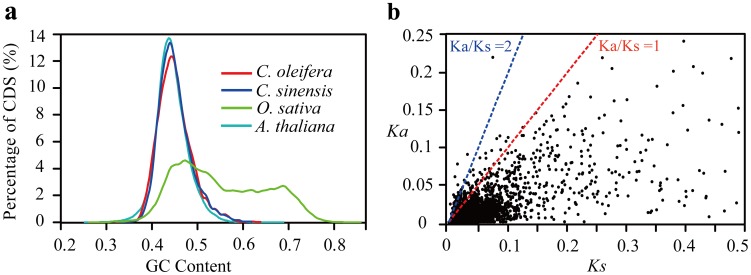
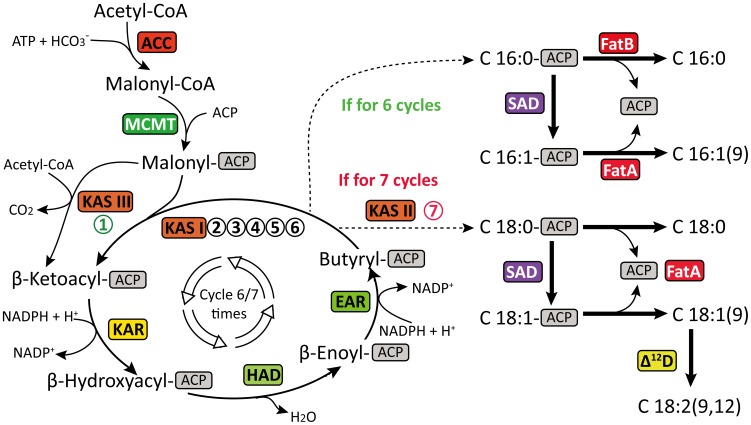
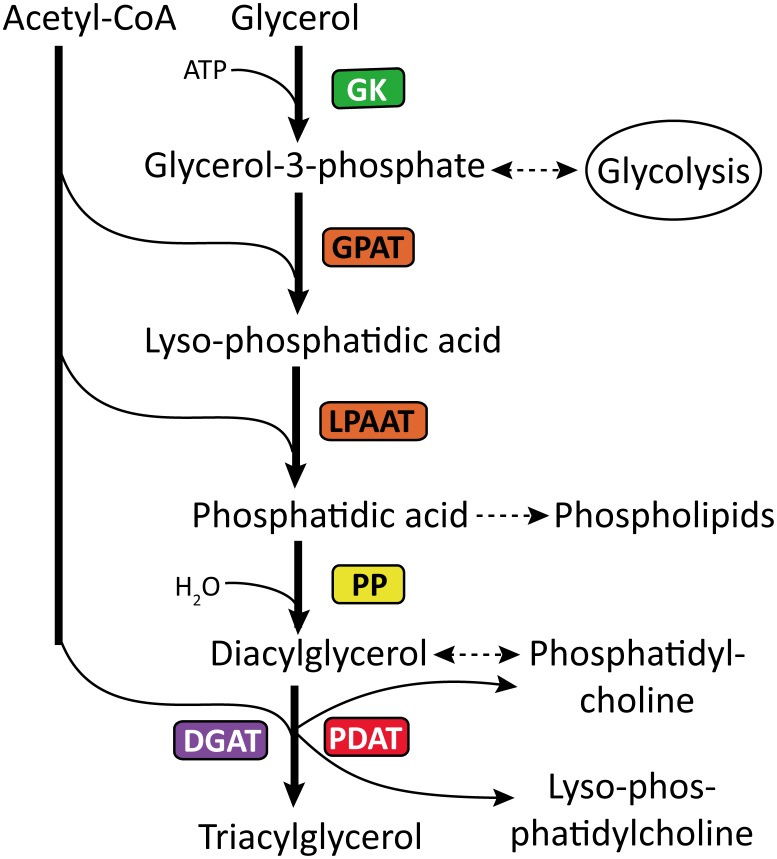
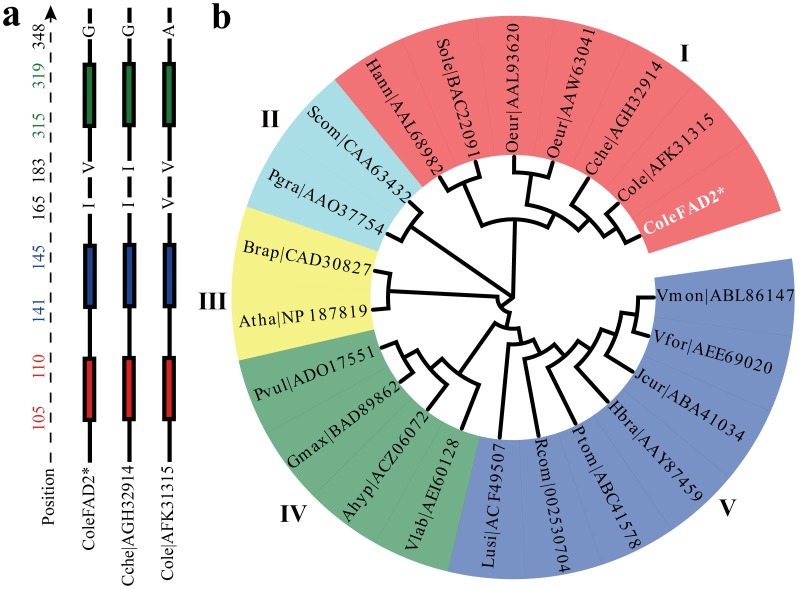
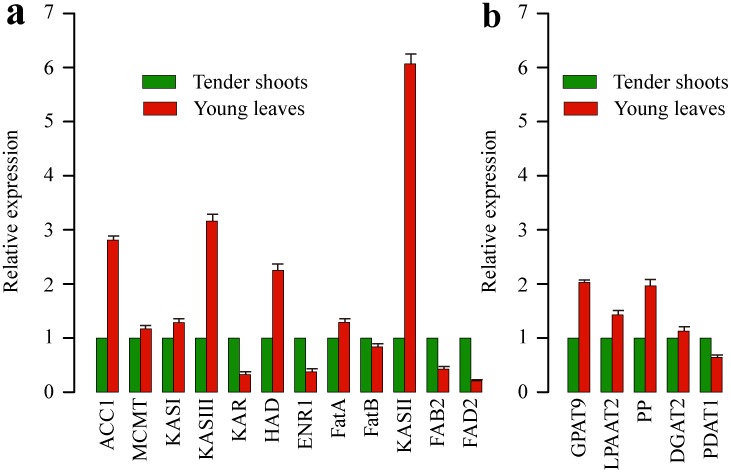
Results: Here, using the 454 GS-FLX sequencing platform, we generated approximately 600,000 RNA-Seq reads from four tissues of C. oleifera. These reads were trimmed and assembled into 104,842 non-redundant putative transcripts with a total length of ∼38.9 Mb, representing more than 218-fold of all the C. oleifera sequences currently deposited in the GenBank (as of March 2014). Based on the BLAST similarity searches, nearly 42.6% transcripts could be annotated with known genes, conserved domains, or Gene Ontology (GO) terms. Comparisons with the cultivated tea tree, C. sinensis, identified 3,022 pairs of orthologs, of which 211 exhibited the evidence under positive selection. Pathway analysis detected the majority of genes potentially related to lipid metabolism. Evolutionary analysis of omega-6 fatty acid desaturase (FAD2) genes among 20 oil-plants unexpectedly suggests that a parallel evolution may occur between C. oleifera and Olea oleifera. Additionally, more than 2,300 simple sequence repeats (SSRs) and 20,200 single-nucleotide polymorphisms (SNPs) were detected in the C. oleifera transcriptome.
Conclusions: The generated transcriptome represents a considerable increase in the number of sequences deposited in the public databases, providing an unprecedented opportunity to discover all related-genes associated with lipid metabolic pathway in C. oleifera. It will greatly enhance the generation of new varieties of C. oleifera with increased yields and high quality.
Conflict of interest statement
Figures


References
-
- Yu Y, Ren S, Tan K (1999) Study on climatic regionalization and layer and belt distribution of oiltea camellia quality in China. J Nat Res 14: 123–127.
-
- Shanan H, Ying G (1982) The comprehensive utilization of camellia fruits. Am Camellia Yearbk 37: 104–107.
-
- Stack L, Ruter J (2006) Teaoil Camellia-Eastern “Olive” for the world. In: XXVII International Horticultural Congress-IHC2006: International Symposium on Asian Plants with Unique Horticultural 769: . pp. 43–48.
-
- Xia L, Zhang A, Xiao T (1993) An introduction to the utilization of camellia oil in China. Am Camellia Yearbk 48: 12–15.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
