

Regulation of male sex determination: genital ridge formation and Sry activation in mice
- PMID: 25139092
- PMCID: PMC4233110
- DOI: 10.1007/s00018-014-1703-3
Regulation of male sex determination: genital ridge formation and Sry activation in mice
Abstract
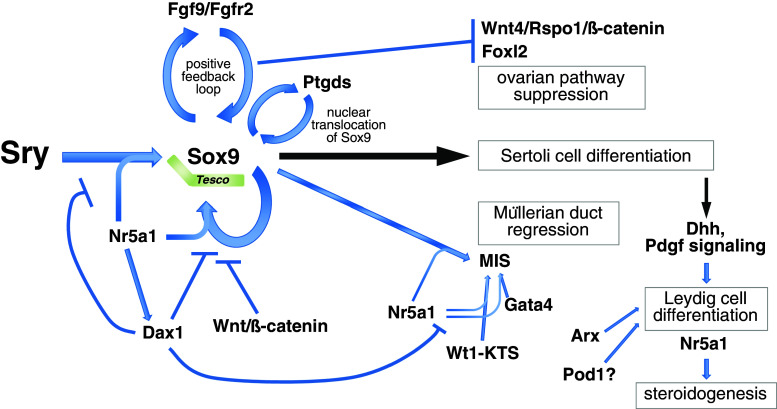
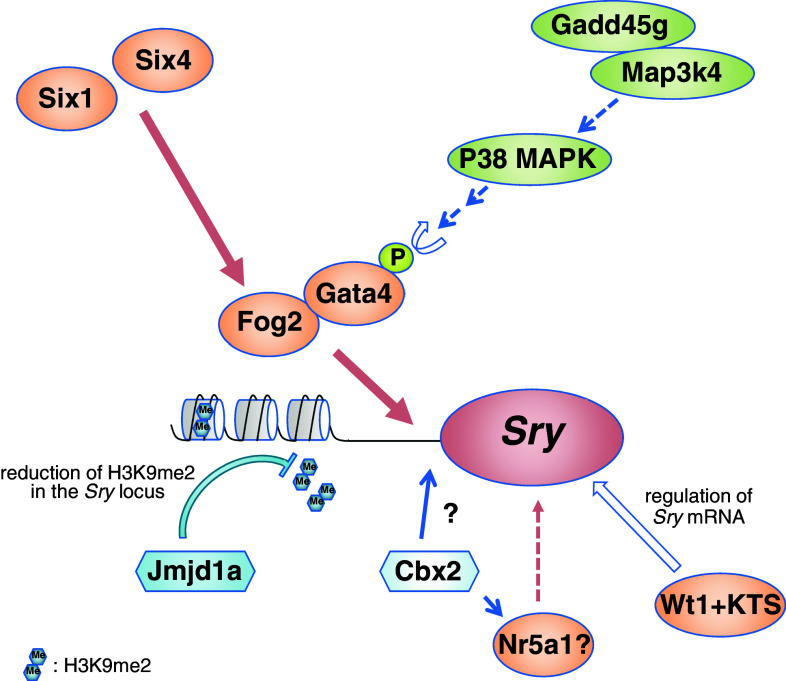
Sex determination is essential for the sexual reproduction to generate the next generation by the formation of functional male or female gametes. In mammals, primary sex determination is commenced by the presence or absence of the Y chromosome, which controls the fate of the gonadal primordium. The somatic precursor of gonads, the genital ridge is formed at the mid-gestation stage and gives rise to one of two organs, a testis or an ovary. The fate of the genital ridge, which is governed by the differentiation of somatic cells into Sertoli cells in the testes or granulosa cells in the ovaries, further determines the sex of an individual and their germ cells. Mutation studies in human patients with disorders of sex development and mouse models have revealed factors that are involved in mammalian sex determination. In most of mammals, a single genetic trigger, the Y-linked gene Sry (sex determination region on Y chromosome), regulates testicular differentiation. Despite identification of Sry in 1990, precise mechanisms underlying the sex determination of bipotential genital ridges are still largely unknown. Here, we review the recent progress that has provided new insights into the mechanisms underlying genital ridge formation as well as the regulation of Sry expression and its functions in male sex determination of mice.
Figures

References
-
- Anderson R, Copeland TK, Scholer H, Heasman J, Wylie C. The onset of germ cell migration in the mouse embryo. Mech Dev. 2000;91:61–68. - PubMed
-
- Ginsburg M, Snow MH, McLaren A. Primordial germ cells in the mouse embryo during gastrulation. Development. 1990;110:521–528. - PubMed
-
- Hara K, Kanai-Azuma M, Uemura M, Shitara H, Taya C, et al. Evidence for crucial role of hindgut expansion in directing proper migration of primordial germ cells in mouse early embryogenesis. Dev Biol. 2009;330:427–439. - PubMed
-
- Lawson KA, Hage WJ. Clonal analysis of the origin of primordial germ cells in the mouse. Ciba Found Symp. 1994;182:68–84. - PubMed
-
- Molyneaux KA, Stallock J, Schaible K, Wylie C. Time-lapse analysis of living mouse germ cell migration. Dev Biol. 2001;240:488–498. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
