

Super-resolution imaging reveals that loss of the C-terminus of connexin43 limits microtubule plus-end capture and NaV1.5 localization at the intercalated disc
- PMID: 25139742
- PMCID: PMC4296112
- DOI: 10.1093/cvr/cvu195
Super-resolution imaging reveals that loss of the C-terminus of connexin43 limits microtubule plus-end capture and NaV1.5 localization at the intercalated disc
Abstract
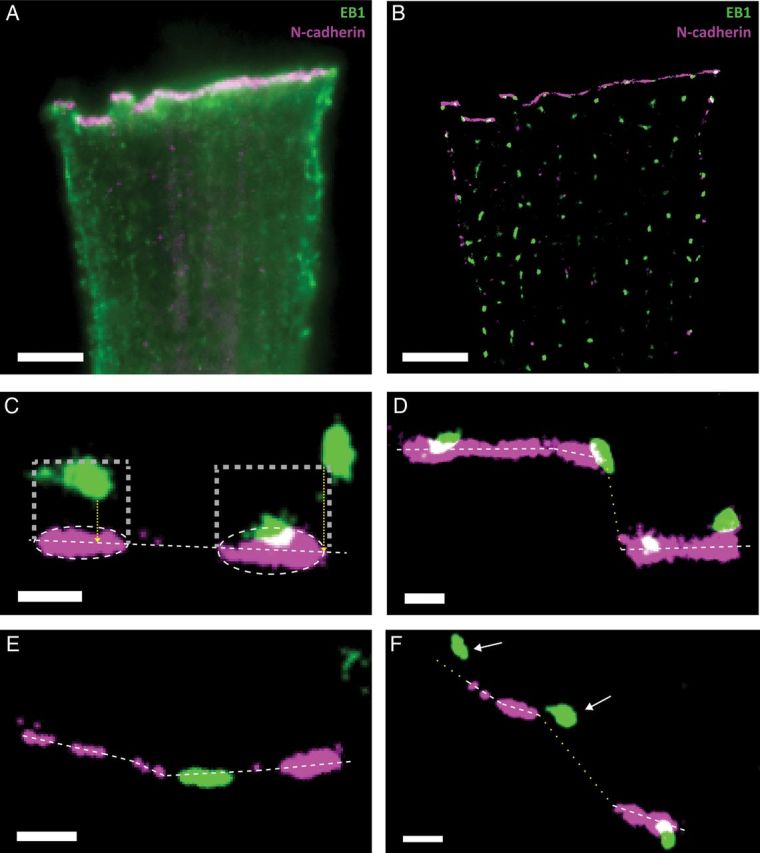
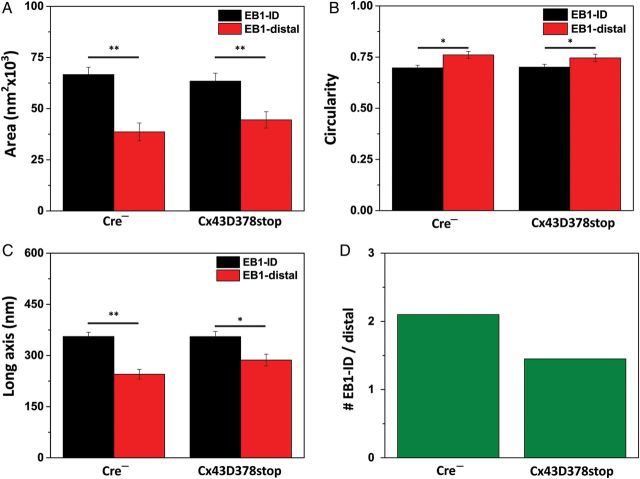
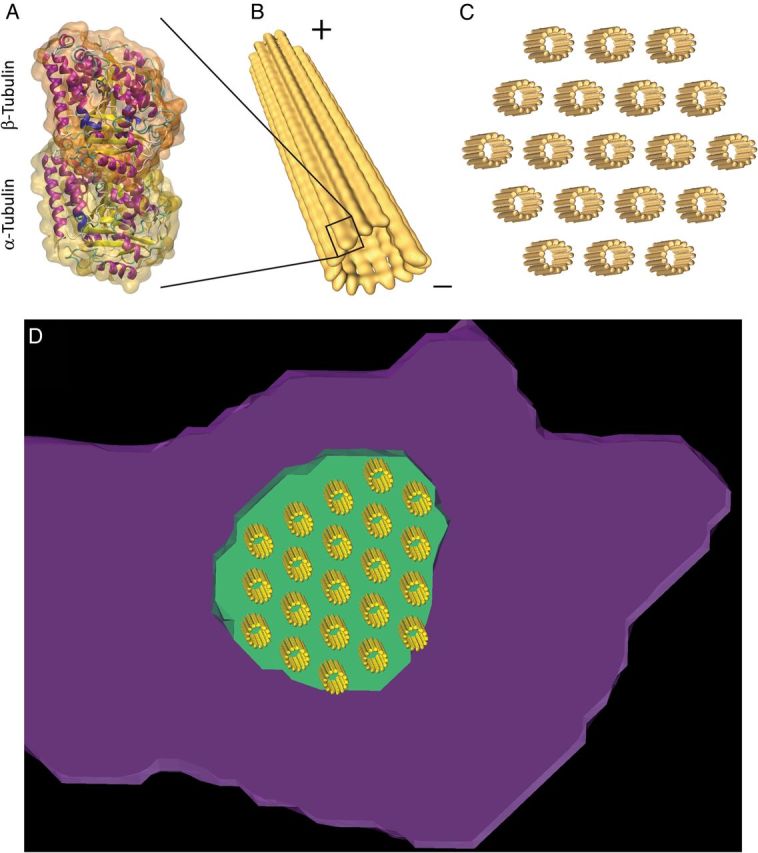
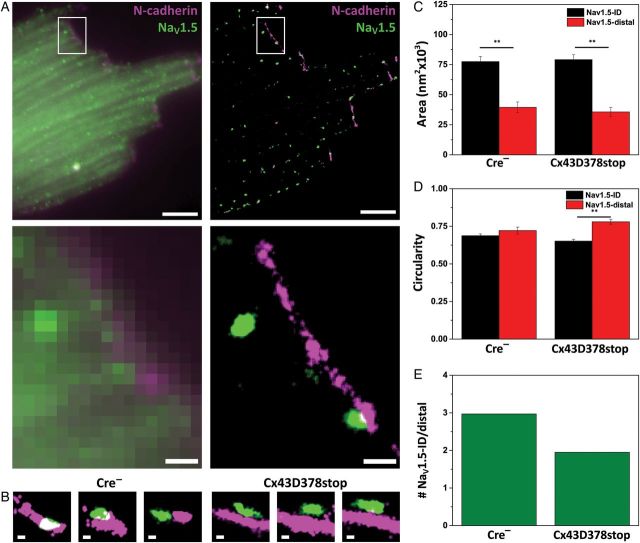
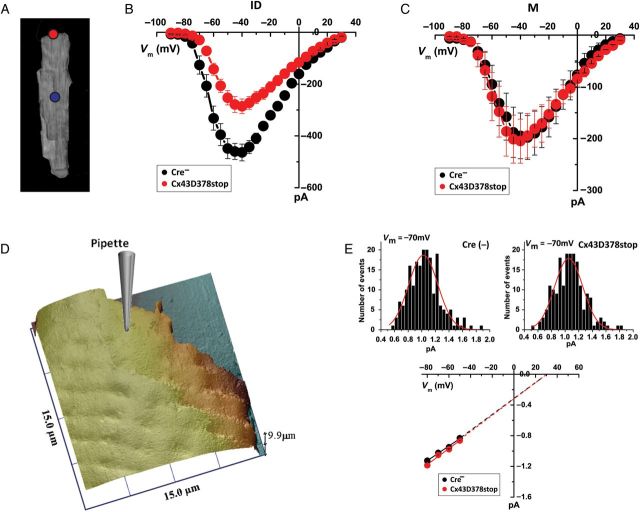
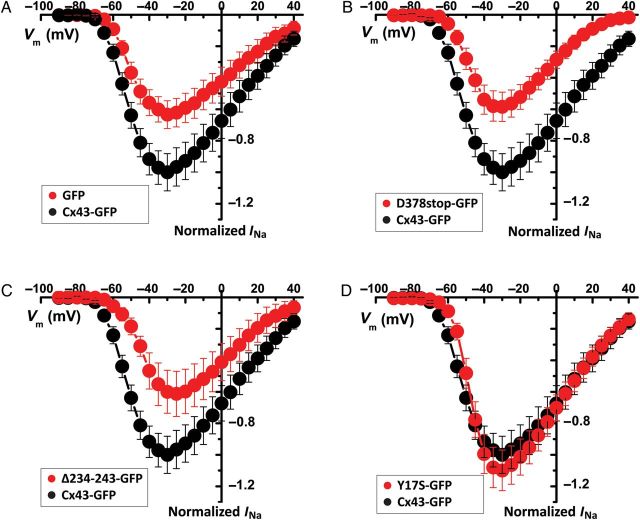
Aims: It is well known that connexin43 (Cx43) forms gap junctions. We recently showed that Cx43 is also part of a protein-interacting network that regulates excitability. Cardiac-specific truncation of Cx43 C-terminus (mutant 'Cx43D378stop') led to lethal arrhythmias. Cx43D378stop localized to the intercalated disc (ID); cell-cell coupling was normal, but there was significant sodium current (INa) loss. We proposed that the microtubule plus-end is at the crux of the Cx43-INa relation. Yet, specific localization of relevant molecular players was prevented due to the resolution limit of fluorescence microscopy. Here, we use nanoscale imaging to establish: (i) the morphology of clusters formed by the microtubule plus-end tracking protein 'end-binding 1' (EB1), (ii) their position, and that of sodium channel alpha-subunit NaV1.5, relative to N-cadherin-rich sites, and (iii) the role of Cx43 C-terminus on the above-mentioned parameters and on the location-specific function of INa.
Methods and results: Super-resolution fluorescence localization microscopy in murine adult cardiomyocytes revealed EB1 and NaV1.5 as distinct clusters preferentially localized to N-cadherin-rich sites. Extent of co-localization decreased in Cx43D378stop cells. Macropatch and scanning patch clamp showed reduced INa exclusively at cell end, without changes in unitary conductance. Experiments in Cx43-modified HL1 cells confirmed the relation between Cx43, INa, and microtubules.
Conclusions: NaV1.5 and EB1 localization at the cell end is Cx43-dependent. Cx43 is part of a molecular complex that determines capture of the microtubule plus-end at the ID, facilitating cargo delivery. These observations link excitability and electrical coupling through a common molecular mechanism.
Keywords: Area composita; Connexome; Cx43; EB1; Microtubule; NaV1.5.
Published on behalf of the European Society of Cardiology. All rights reserved. © The Author 2014. For permissions please email: journals.permissions@oup.com.
Figures

Comment in
-
Elucidating sodium channel NaV1.5 clustering in cardiac myocytes using super-resolution techniques.Cardiovasc Res. 2014 Nov 1;104(2):231-3. doi: 10.1093/cvr/cvu221. Epub 2014 Oct 23. Cardiovasc Res. 2014. PMID: 25341893 No abstract available.
References
-
- Akar FG, Nass RD, Hahn S, Cingolani E, Shah M, Hesketh GG, DiSilvestre D, Tunin RS, Kass DA, Tomaselli GF. Dynamic changes in conduction velocity and gap junction properties during development of pacing-induced heart failure. Am J Physiol Heart Circ Physiol. 2007;293:H1223–H1230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
