

Scrib:Rac1 interactions are required for the morphogenesis of the ventricular myocardium
- PMID: 25139745
- PMCID: PMC4174891
- DOI: 10.1093/cvr/cvu193
Scrib:Rac1 interactions are required for the morphogenesis of the ventricular myocardium
Abstract
Aims: The organization and maturation of ventricular cardiomyocytes from the embryonic to the adult form is crucial for normal cardiac function. We have shown that a polarity protein, Scrib, may be involved in regulating the early stages of this process. Our goal was to establish whether Scrib plays a cell autonomous role in the ventricular myocardium, and whether this involves well-known polarity pathways.
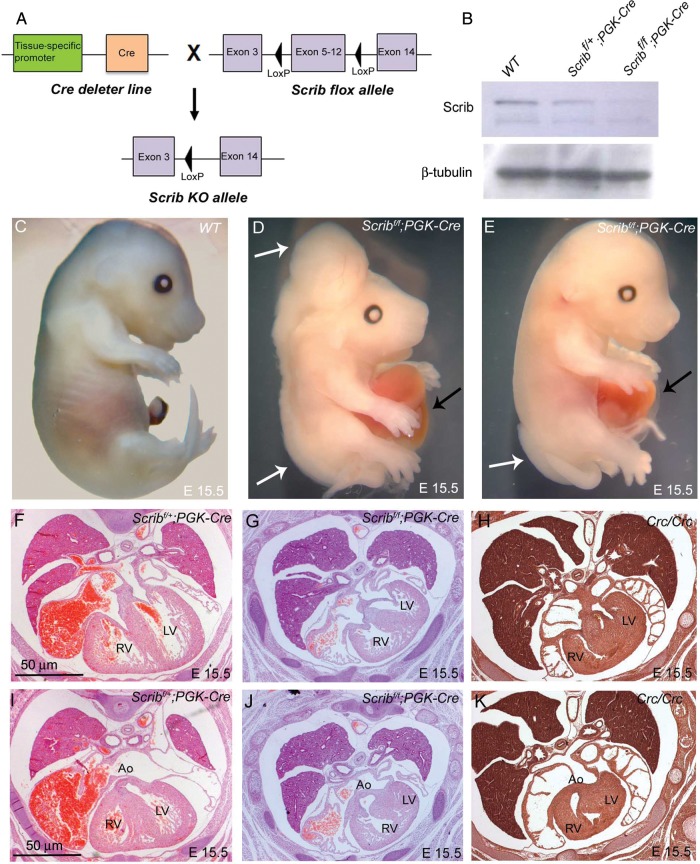
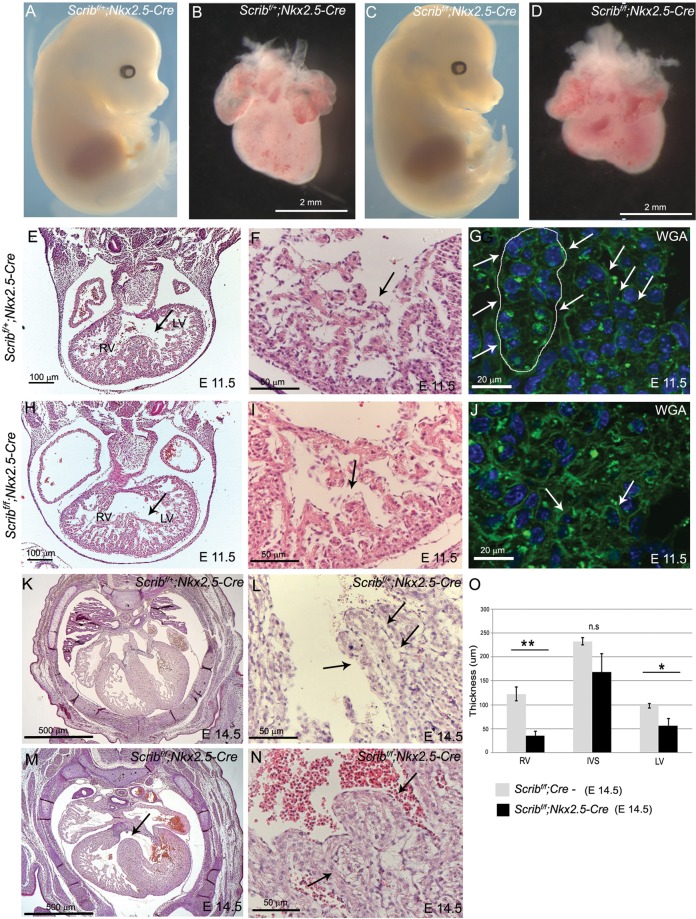
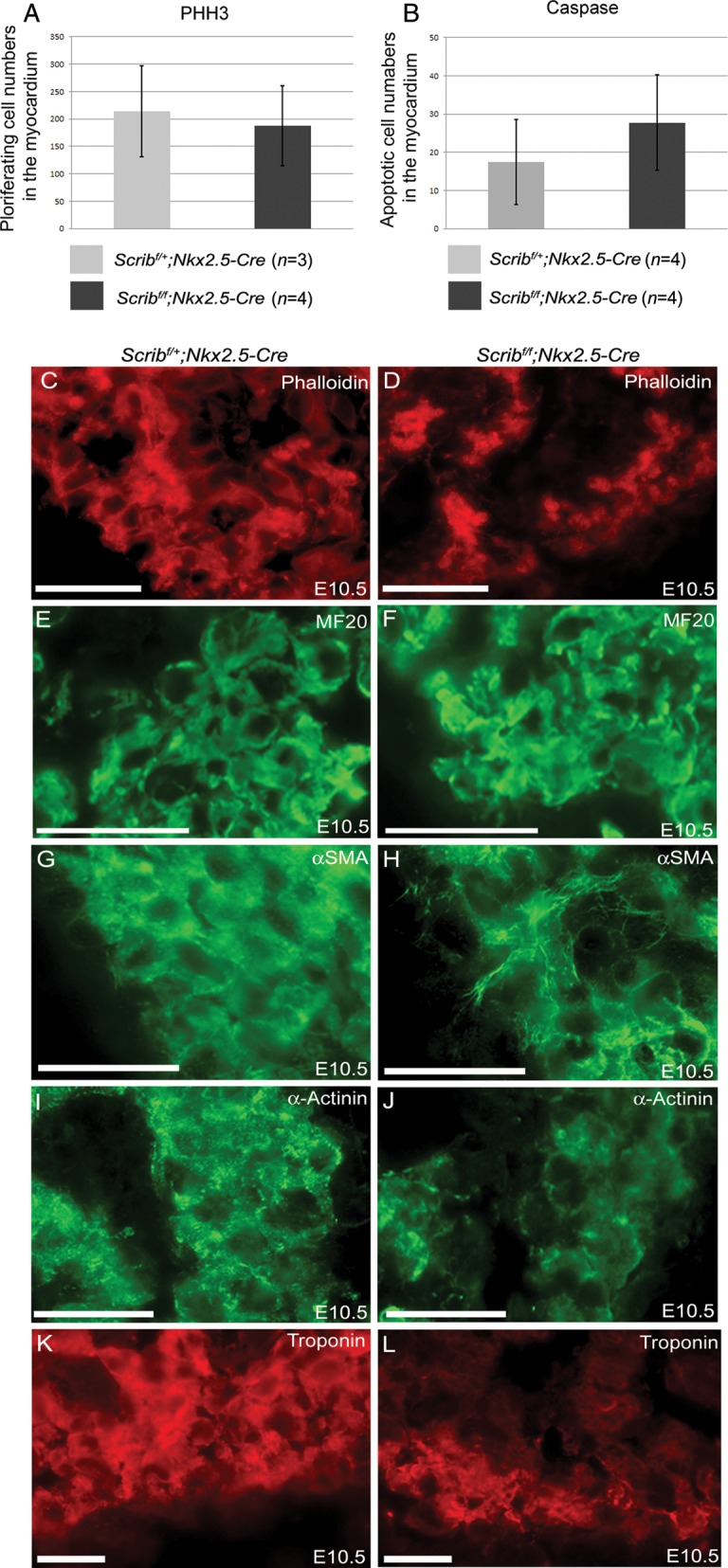
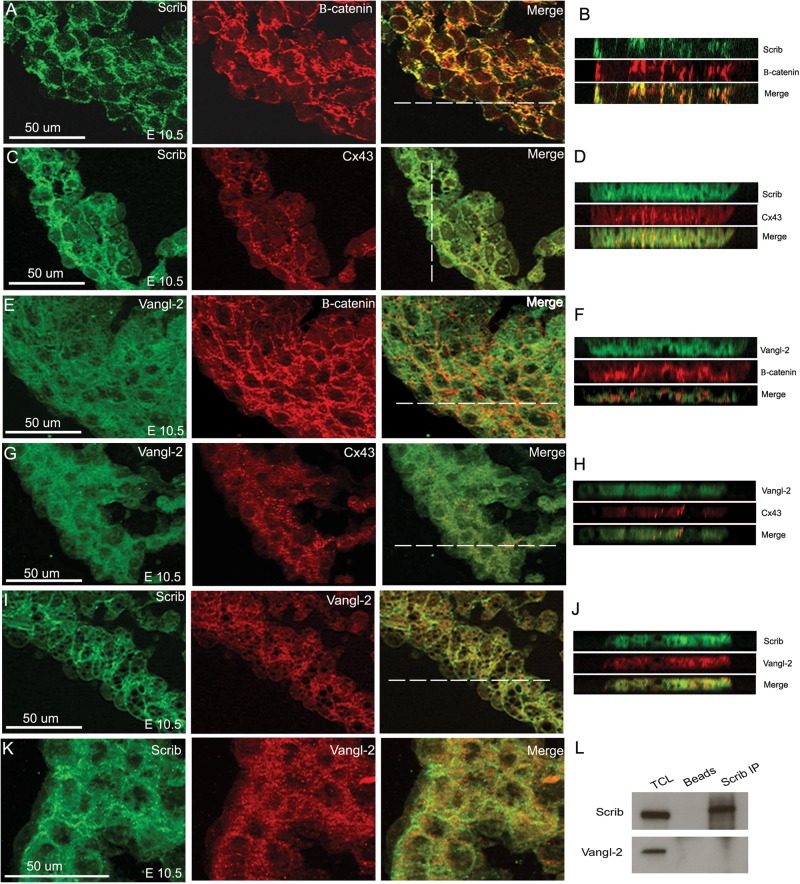
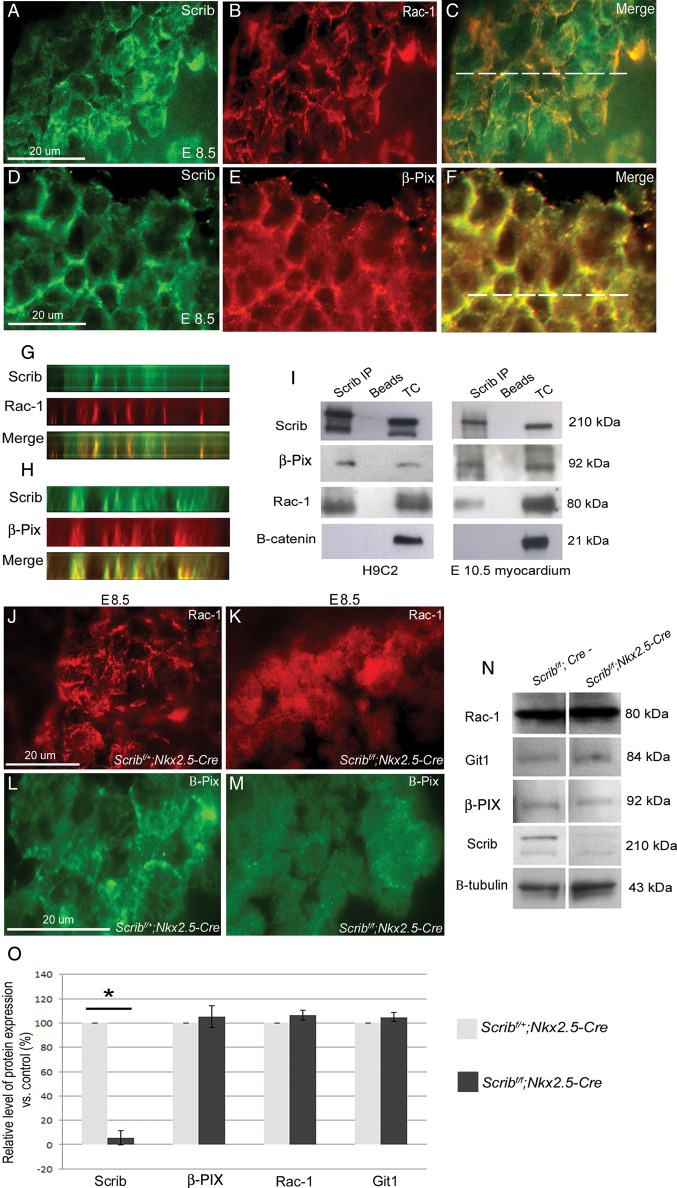
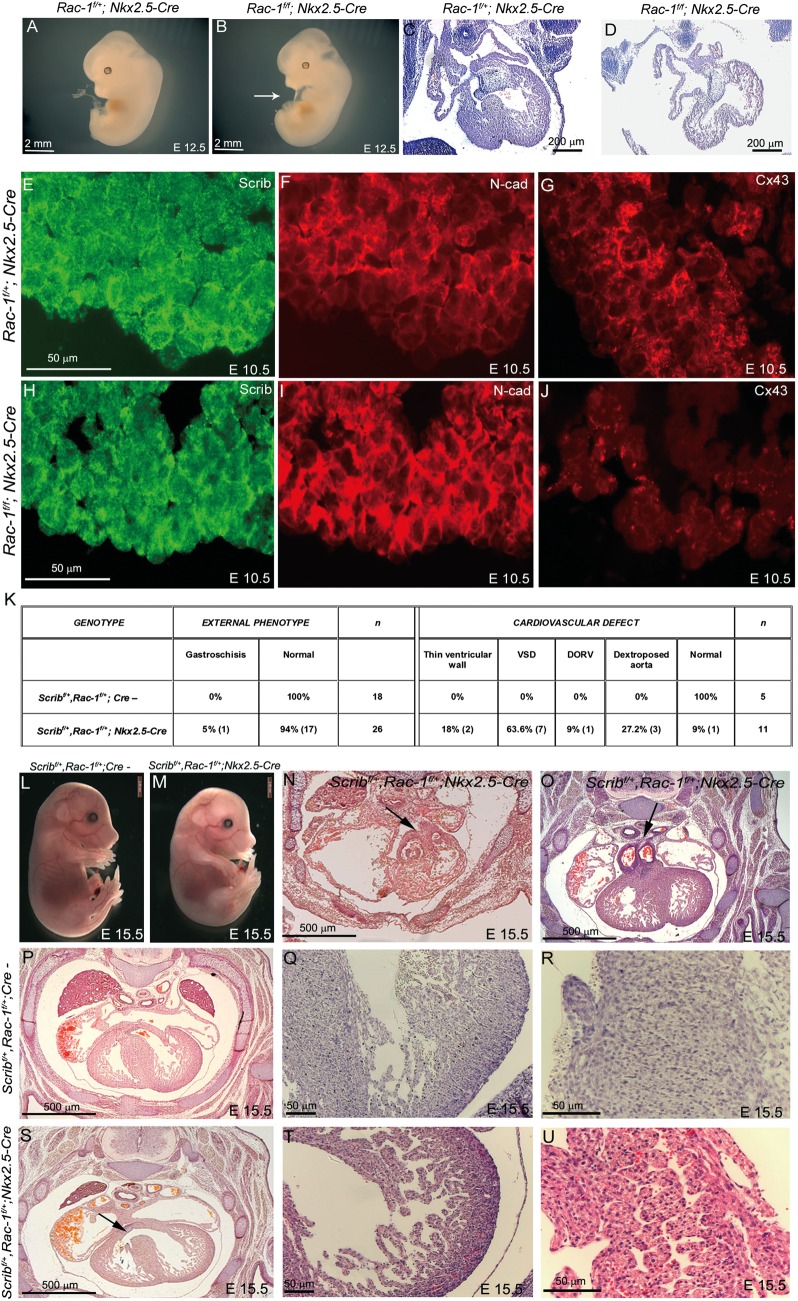
Methods and results: Deletion of Scrib in cardiac precursors utilizing Scrib(flox) mice together with the Nkx2.5-Cre driver resulted in disruption of the cytoarchitecture of the forming trabeculae and ventricular septal defects. Although the majority of mice lacking Scrib in the myocardium survived to adulthood, they developed marked cardiac fibrosis. Scrib did not physically interact with the planar cell polarity (PCP) protein, Vangl2, in early cardiomyocytes as it does in other tissues, suggesting that the anomalies did not result from disruption of PCP signalling. However, Scrib interacted with Rac1 physically in embryonic cardiomyocytes and genetically to result in ventricular abnormalities, suggesting that this interaction is crucial for the development of the early myocardium.
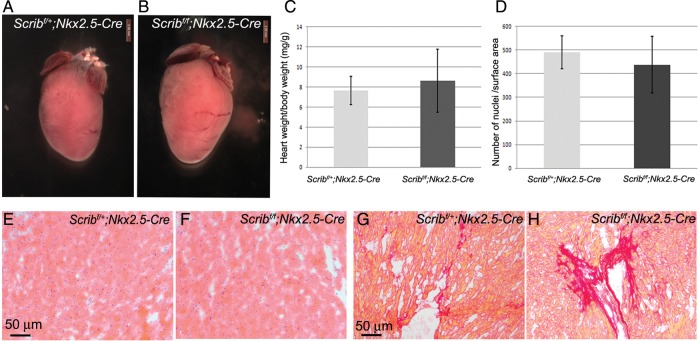
Conclusions: The Scrib-Rac1 interaction plays a crucial role in the organization of developing cardiomyocytes and formation of the ventricular myocardium. Thus, we have identified a novel signalling pathway in the early, functioning, heart muscle. These data also show that the foetus can recover from relatively severe abnormalities in prenatal ventricular development, although cardiac fibrosis can be a long-term consequence.
Keywords: Cardiac development; Cardiomyocytes; Polarity; Rac1; Scrib; Ventricular myocardium.
© The Author 2014. Published by Oxford University Press on behalf of the European Society of Cardiology.
Figures
References
-
- Anderson RH, Ho SY, Redmann K, Sanchez-Quintana D, Lunkenheimer PP. The anatomical arrangement of the myocardial cells making up the ventricular mass. Eur J Cardiothorac Surg. 2005;28:517–525. - PubMed
-
- Olivotto I, Cecchi F, Poggesi C, Yacoub MH. Developmental origins of hypertrophic cardiomyopathy phenotypes: a unifying hypothesis. Nat Rev Cardiol. 2009;6:317–321. - PubMed
-
- Assémat E, Bazellières E, Pallesi-Pocachard E, Le Bivic A, Massey-Harroche D. Polarity complex proteins. Biochim Biophys Acta. 2008;1778:614–630. - PubMed
-
- Elsum I, Yates L, Humbert PO, Richardson HE. The Scribble-Dlg-Lgl polarity module in development and cancer: from flies to man. Essays Biochem. 2012;53:141. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
