

Annexin A2 mediates Mycoplasma pneumoniae community-acquired respiratory distress syndrome toxin binding to eukaryotic cells
- PMID: 25139904
- PMCID: PMC4147866
- DOI: 10.1128/mBio.01497-14
Annexin A2 mediates Mycoplasma pneumoniae community-acquired respiratory distress syndrome toxin binding to eukaryotic cells
Abstract
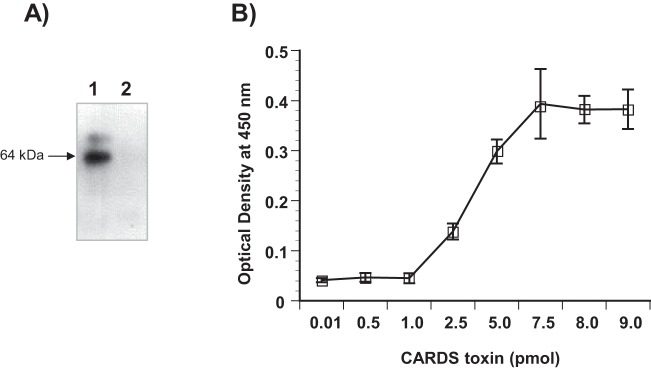
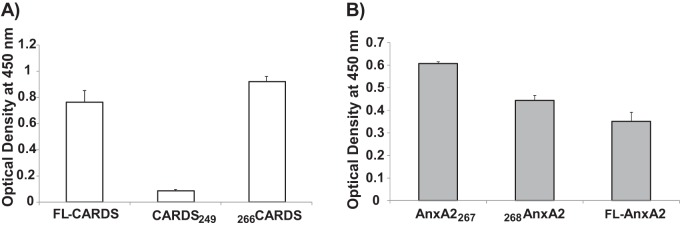
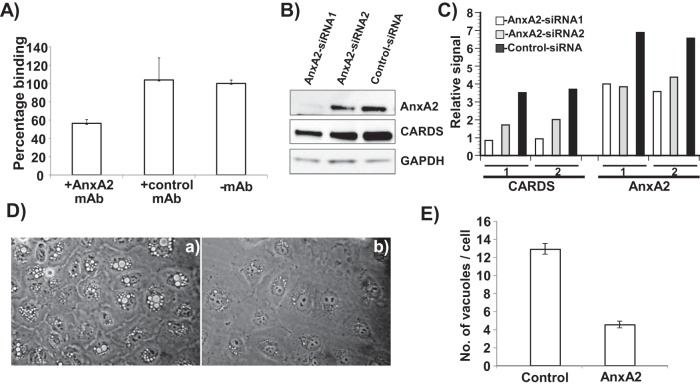
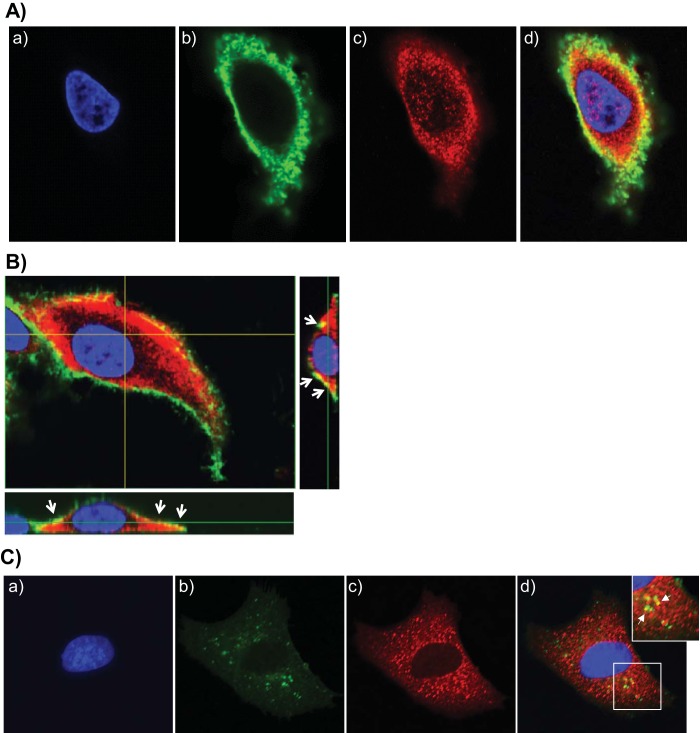
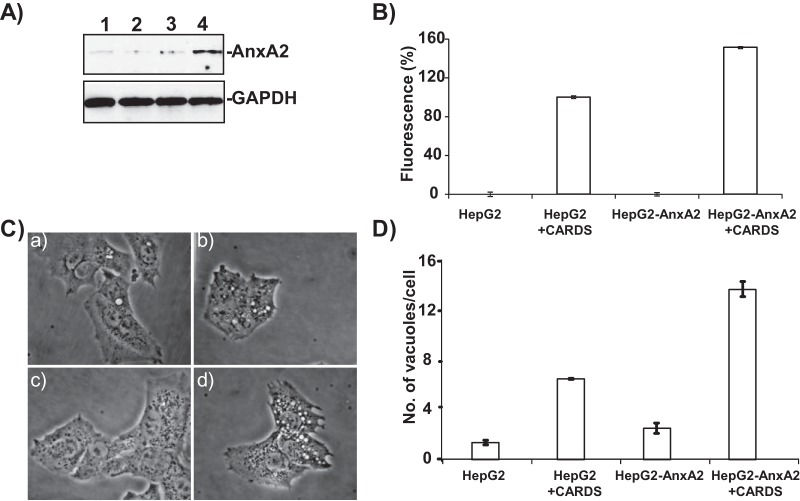
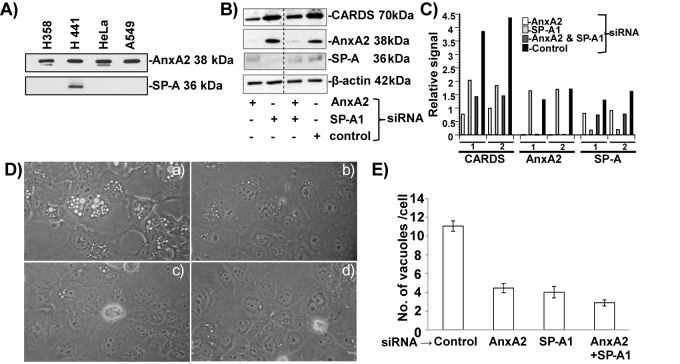
Mycoplasma pneumoniae synthesizes a novel human surfactant protein A (SP-A)-binding cytotoxin, designated community-acquired respiratory distress syndrome (CARDS) toxin, that exhibits ADP-ribosylating and vacuolating activities in mammalian cells and is directly linked to a range of acute and chronic airway diseases, including asthma. In our attempt to detect additional CARDS toxin-binding proteins, we subjected the membrane fraction of human A549 airway cells to affinity chromatography using recombinant CARDS toxin as bait. A 36-kDa A549 cell membrane protein bound to CARDS toxin and was identified by time of flight (TOF) mass spectroscopy as annexin A2 (AnxA2) and verified by immunoblotting with anti-AnxA2 monoclonal antibody. Dose-dependent binding of CARDS toxin to recombinant AnxA2 reinforced the specificity of the interaction, and further studies revealed that the carboxy terminus of CARDS toxin mediated binding to AnxA2. In addition, pretreatment of viable A549 cells with anti-AnxA2 monoclonal antibody or AnxA2 small interfering RNA (siRNA) reduced toxin binding and internalization. Immunofluorescence analysis of CARDS toxin-treated A549 cells demonstrated the colocalization of CARDS toxin with cell surface-associated AnxA2 upon initial binding and with intracellular AnxA2 following toxin internalization. HepG2 cells, which express low levels of AnxA2, were transfected with a plasmid expressing AnxA2 protein, resulting in enhanced binding of CARDS toxin and increased vacuolization. In addition, NCI-H441 cells, which express both AnxA2 and SP-A, upon AnxA2 siRNA transfection, showed decreased binding and subsequent vacuolization. These results indicate that CARDS toxin recognizes AnxA2 as a functional receptor, leading to CARDS toxin-induced changes in mammalian cells.
Importance: Host cell susceptibility to bacterial toxins is usually determined by the presence and abundance of appropriate receptors, which provides a molecular basis for toxin target cell specificities. To perform its ADP-ribosylating and vacuolating activities, community-acquired respiratory distress syndrome (CARDS) toxin must bind to host cell surfaces via receptor-mediated events in order to be internalized and trafficked effectively. Earlier, we reported the binding of CARDS toxin to surfactant protein A (SP-A), and here we show how CARDS toxin uses an alternative receptor to execute its pathogenic properties. CARDS toxin binds selectively to annexin A2 (AnxA2), which exists both on the cell surface and intracellularly. Since AnxA2 regulates membrane dynamics at early stages of endocytosis and trafficking, it serves as a distinct receptor for CARDS toxin binding and internalization and enhances CARDS toxin-induced vacuolization in mammalian cells.
Copyright © 2014 Somarajan et al.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
