

Phospholipase A2 regulates eicosanoid class switching during inflammasome activation
- PMID: 25139986
- PMCID: PMC4156727
- DOI: 10.1073/pnas.1404372111
Phospholipase A2 regulates eicosanoid class switching during inflammasome activation
Abstract
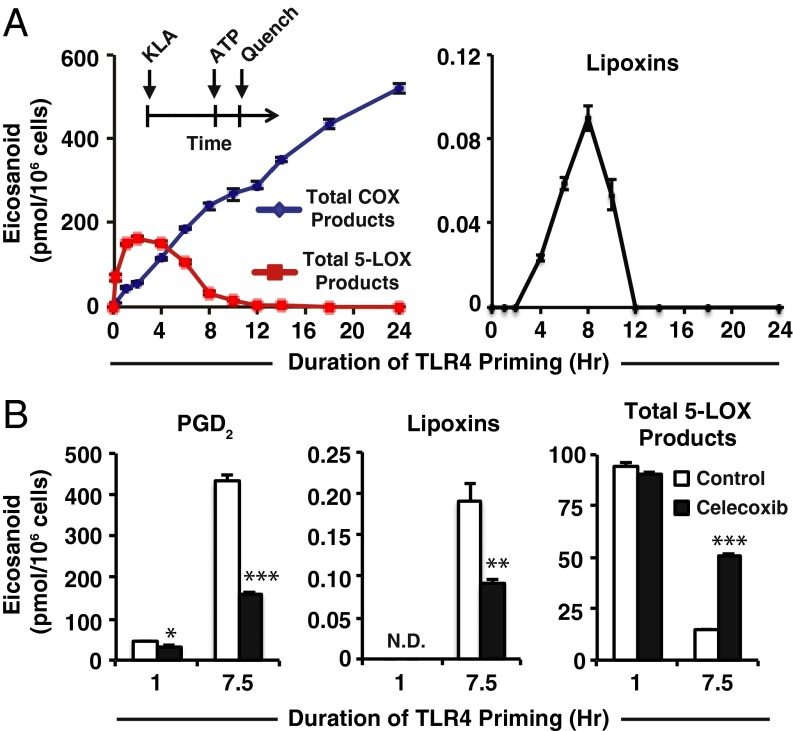
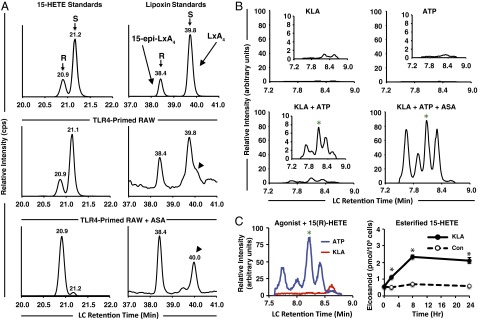
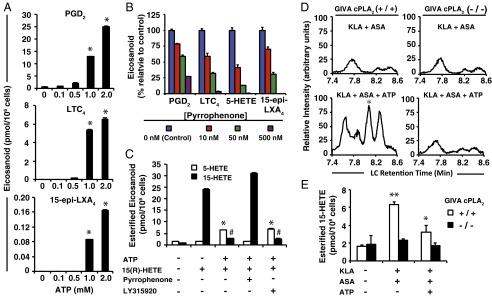
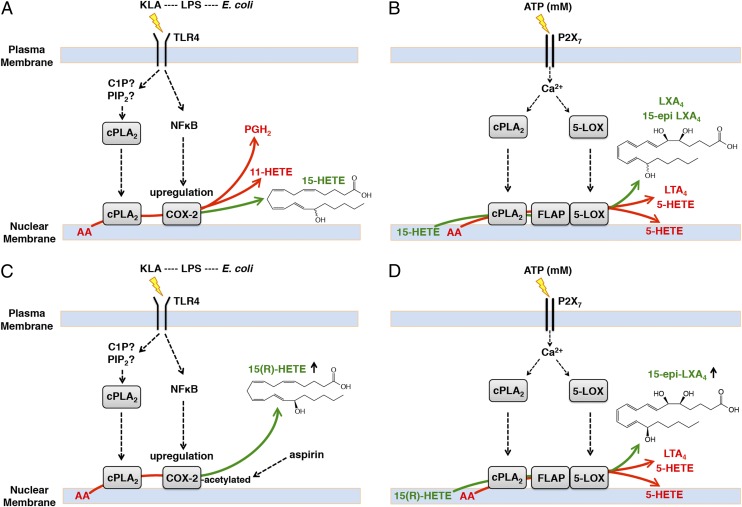
Initiation and resolution of inflammation are considered to be tightly connected processes. Lipoxins (LX) are proresolution lipid mediators that inhibit phlogistic neutrophil recruitment and promote wound-healing macrophage recruitment in humans via potent and specific signaling through the LXA4 receptor (ALX). One model of lipoxin biosynthesis involves sequential metabolism of arachidonic acid by two cell types expressing a combined transcellular metabolon. It is currently unclear how lipoxins are efficiently formed from precursors or if they are directly generated after receptor-mediated inflammatory commitment. Here, we provide evidence for a pathway by which lipoxins are generated in macrophages as a consequence of sequential activation of toll-like receptor 4 (TLR4), a receptor for endotoxin, and P2X7, a purinergic receptor for extracellular ATP. Initial activation of TLR4 results in accumulation of the cyclooxygenase-2-derived lipoxin precursor 15-hydroxyeicosatetraenoic acid (15-HETE) in esterified form within membrane phospholipids, which can be enhanced by aspirin (ASA) treatment. Subsequent activation of P2X7 results in efficient hydrolysis of 15-HETE from membrane phospholipids by group IVA cytosolic phospholipase A2, and its conversion to bioactive lipoxins by 5-lipoxygenase. Our results demonstrate how a single immune cell can store a proresolving lipid precursor and then release it for bioactive maturation and secretion, conceptually similar to the production and inflammasome-dependent maturation of the proinflammatory IL-1 family cytokines. These findings provide evidence for receptor-specific and combinatorial control of pro- and anti-inflammatory eicosanoid biosynthesis, and potential avenues to modulate inflammatory indices without inhibiting downstream eicosanoid pathways.
Keywords: enzyme coupling; lipidomics; membrane remodeling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ferrari D, et al. The P2X7 receptor: A key player in IL-1 processing and release. J Immunol. 2006;176(7):3877–3883. - PubMed
-
- Ferrari D, et al. Extracellular ATP triggers IL-1 beta release by activating the purinergic P2Z receptor of human macrophages. J Immunol. 1997;159(3):1451–1458. - PubMed
-
- Solle M, et al. Altered cytokine production in mice lacking P2X(7) receptors. J Biol Chem. 2001;276(1):125–132. - PubMed
-
- Kahlenberg JM, Lundberg KC, Kertesy SB, Qu Y, Dubyak GR. Potentiation of caspase-1 activation by the P2X7 receptor is dependent on TLR signals and requires NF-kappaB-driven protein synthesis. J Immunol. 2005;175(11):7611–7622. - PubMed
-
- Suzuki K, et al. Lipopolysaccharide primes human alveolar macrophages for enhanced release of superoxide anion and leukotriene B4: Self-limitations of the priming response with protein synthesis. Am J Respir Cell Mol Biol. 1993;8(5):500–508. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
