

Structure activity relationship of phenolic acid inhibitors of α-synuclein fibril formation and toxicity
- PMID: 25140150
- PMCID: PMC4122169
- DOI: 10.3389/fnagi.2014.00197
Structure activity relationship of phenolic acid inhibitors of α-synuclein fibril formation and toxicity
Abstract
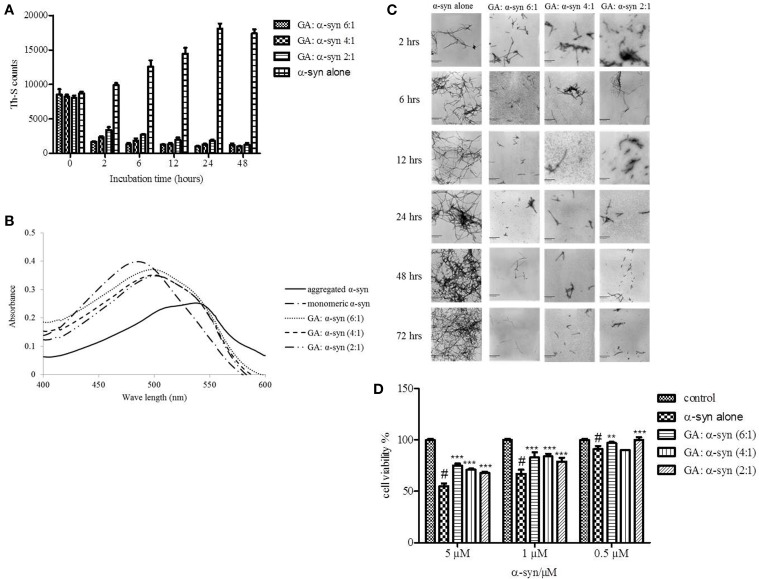
The aggregation of α-synuclein (α-syn) is considered the key pathogenic event in many neurological disorders such as Parkinson's disease (PD), dementia with Lewy bodies and multiple system atrophy, giving rise to a whole category of neurodegenerative diseases known as synucleinopathies. Although the molecular basis of α-syn toxicity has not been precisely elucidated, a great deal of effort has been put into identifying compounds that could inhibit or even reverse the aggregation process. Previous reports indicated that many phenolic compounds are potent inhibitors of α-syn aggregation. The aim of the present study was to assess the anti-aggregating effect of gallic acid (GA) (3,4,5-trihydroxybenzoic acid), a benzoic acid derivative that belongs to a group of phenolic compounds known as phenolic acids. By employing an array of biophysical and biochemical techniques and a cell-viability assay, GA was shown not only to inhibit α-syn fibrillation and toxicity but also to disaggregate preformed α-syn amyloid fibrils. Interestingly, GA was found to bind to soluble, non-toxic oligomers with no β-sheet content, and to stabilize their structure. The binding of GA to the oligomers may represent a potential mechanism of action. Additionally, by using structure activity relationship data obtained from fourteen structurally similar benzoic acid derivatives, it was determined that the inhibition of α-syn fibrillation by GA is related to the number of hydroxyl moieties and their position on the phenyl ring. GA may represent the starting point for designing new molecules that could be used for the treatment of PD and related disorders.
Keywords: Parkinson's disease; aggregation; amyloid fibrils; drug discovery; gallic acid; α-synuclein.
Figures







References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous