

Loss of γ-cytoplasmic actin triggers myofibroblast transition of human epithelial cells
- PMID: 25143399
- PMCID: PMC4196865
- DOI: 10.1091/mbc.E14-03-0815
Loss of γ-cytoplasmic actin triggers myofibroblast transition of human epithelial cells
Abstract
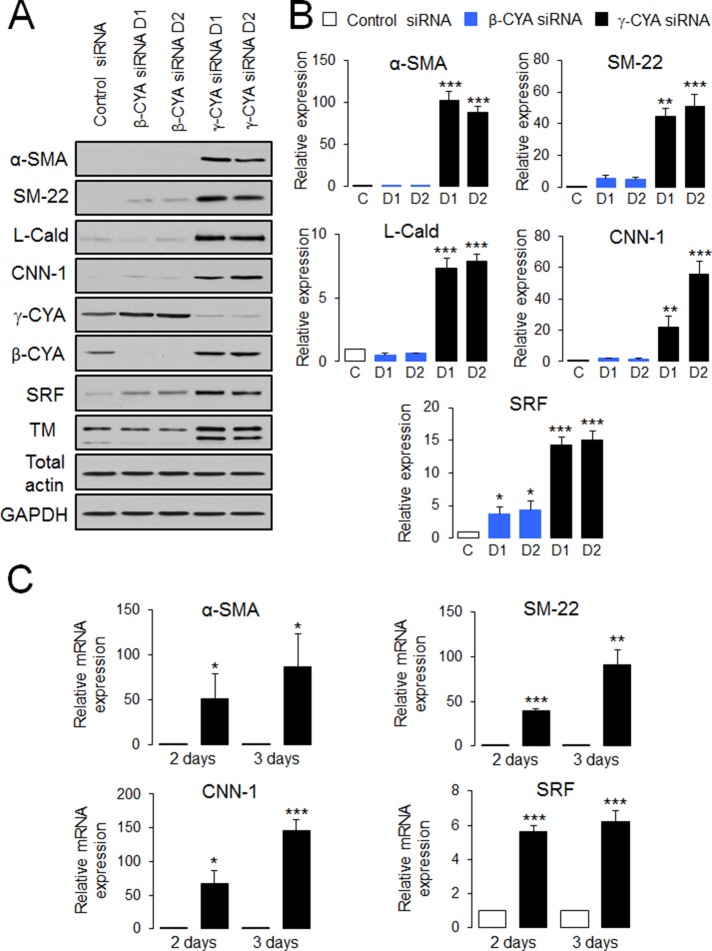
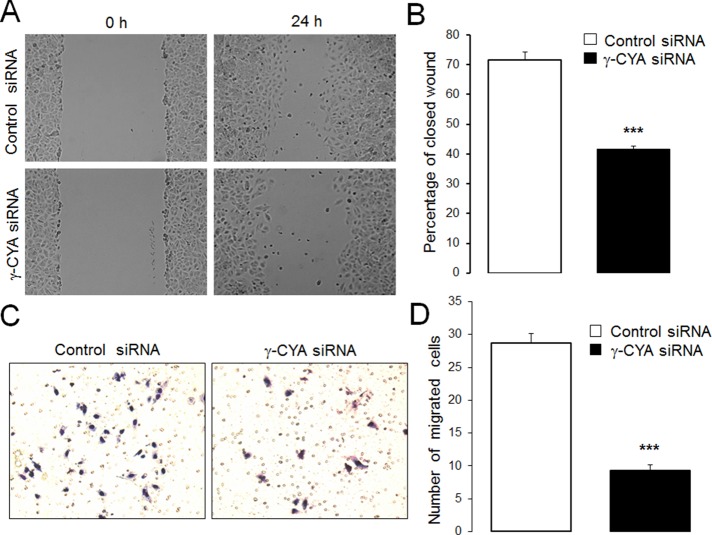
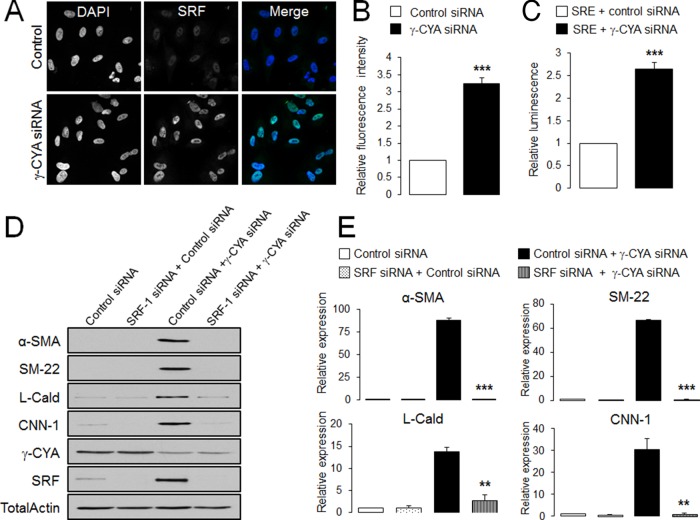
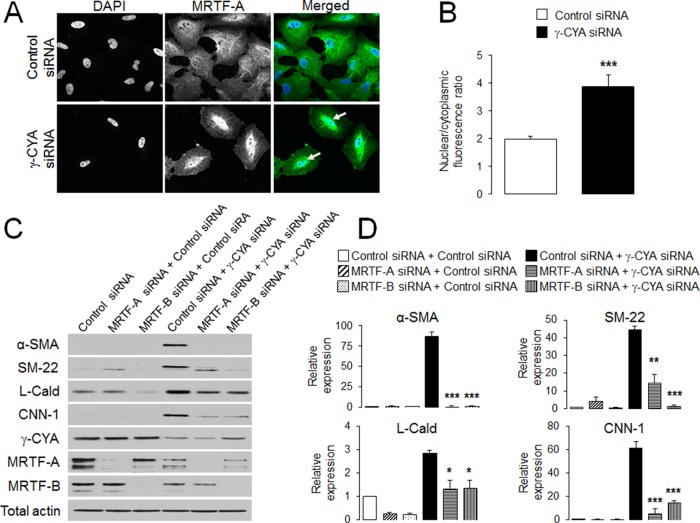
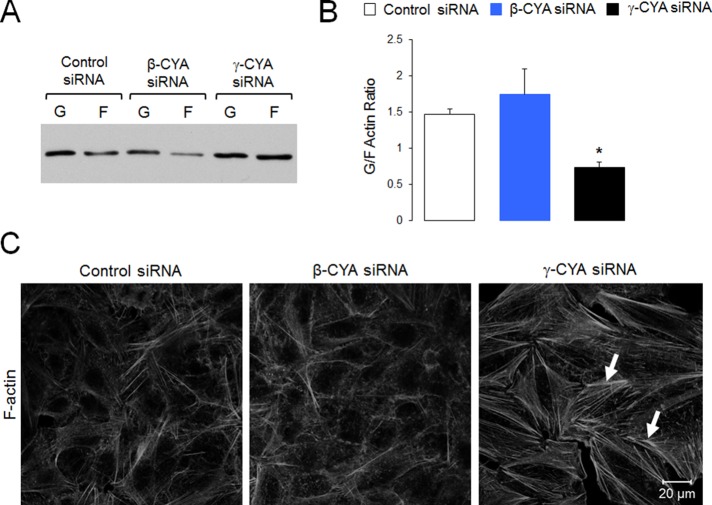
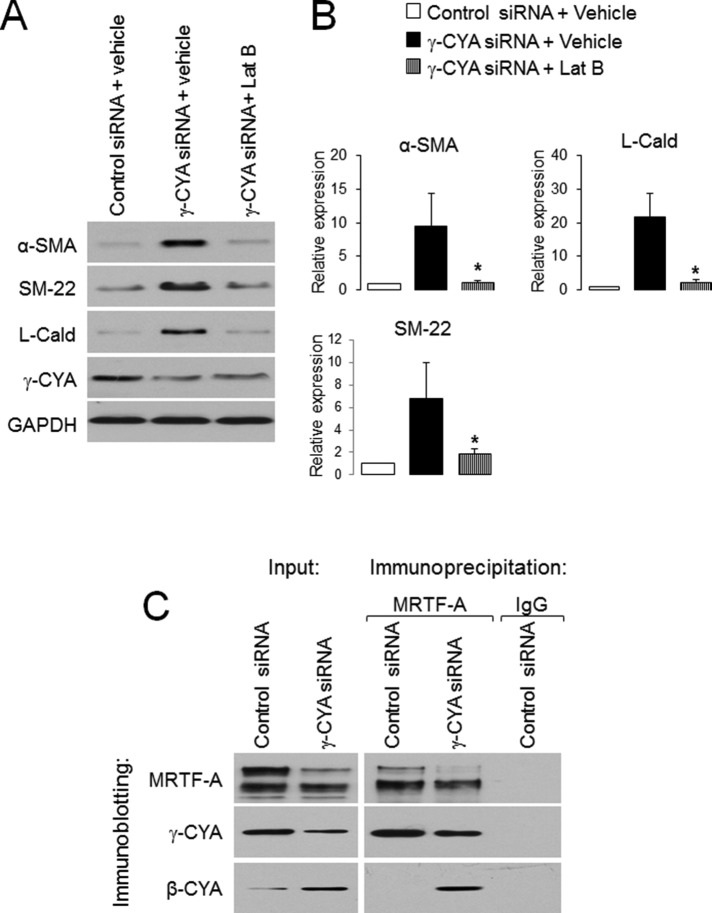
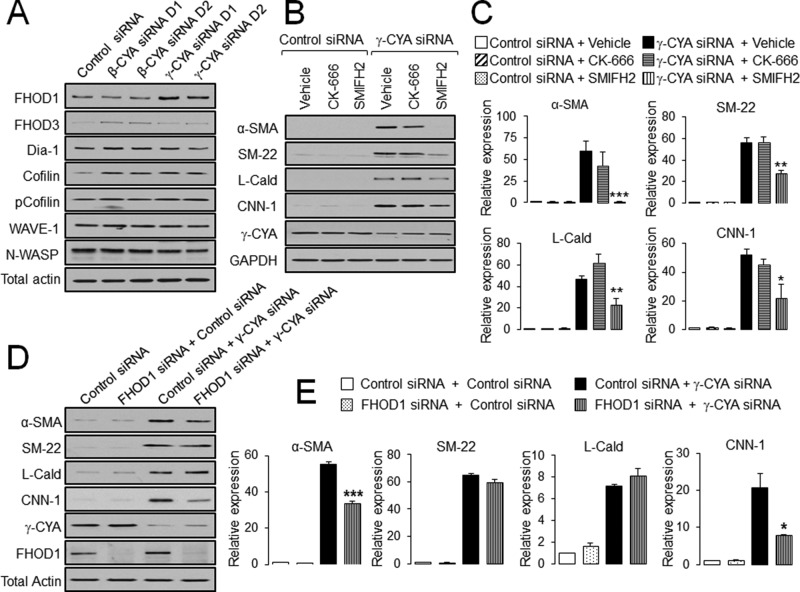
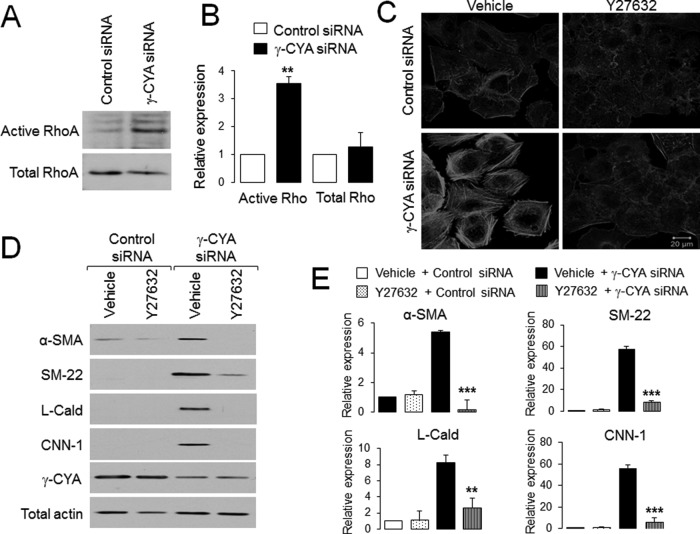
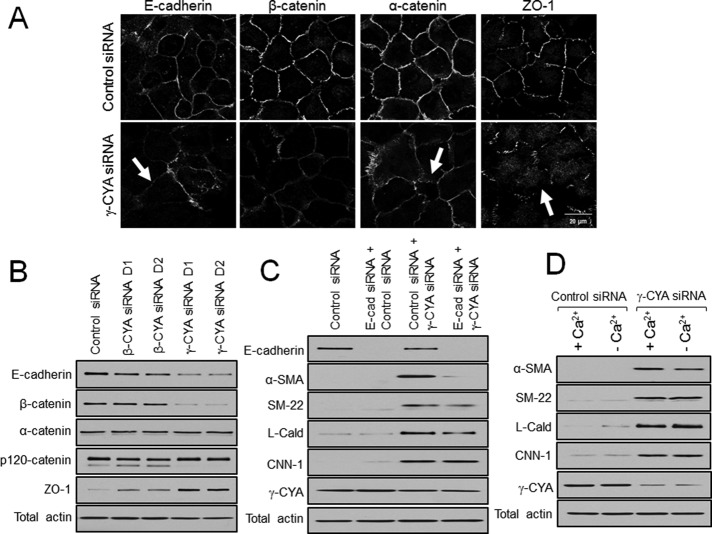
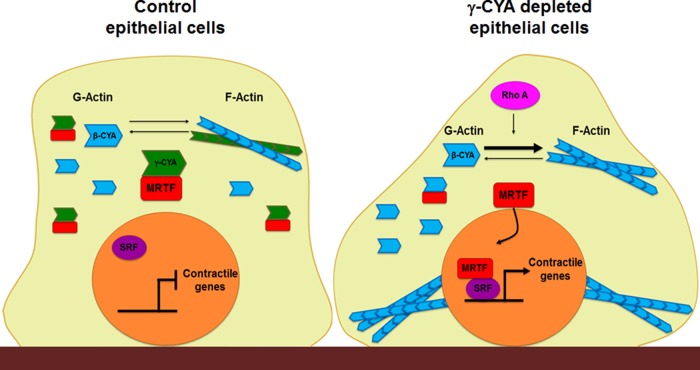
Transdifferentiation of epithelial cells into mesenchymal cells and myofibroblasts plays an important role in tumor progression and tissue fibrosis. Such epithelial plasticity is accompanied by dramatic reorganizations of the actin cytoskeleton, although mechanisms underlying cytoskeletal effects on epithelial transdifferentiation remain poorly understood. In the present study, we observed that selective siRNA-mediated knockdown of γ-cytoplasmic actin (γ-CYA), but not β-cytoplasmic actin, induced epithelial-to-myofibroblast transition (EMyT) of different epithelial cells. The EMyT manifested by increased expression of α-smooth muscle actin and other contractile proteins, along with inhibition of genes responsible for cell proliferation. Induction of EMyT in γ-CYA-depleted cells depended on activation of serum response factor and its cofactors, myocardial-related transcriptional factors A and B. Loss of γ-CYA stimulated formin-mediated actin polymerization and activation of Rho GTPase, which appear to be essential for EMyT induction. Our findings demonstrate a previously unanticipated, unique role of γ-CYA in regulating epithelial phenotype and suppression of EMyT that may be essential for cell differentiation and tissue fibrosis.
© 2014 Lechuga, Baranwal, et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Baarlink C, Wang H, Grosse R. Nuclear actin network assembly by formins regulates the SRF coactivator MAL. Science. 2013;340:864–867. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
