

Atheroprotective immunization with malondialdehyde-modified LDL is hapten specific and dependent on advanced MDA adducts: implications for development of an atheroprotective vaccine
- PMID: 25143462
- PMCID: PMC4174006
- DOI: 10.1194/jlr.M053256
Atheroprotective immunization with malondialdehyde-modified LDL is hapten specific and dependent on advanced MDA adducts: implications for development of an atheroprotective vaccine
Abstract
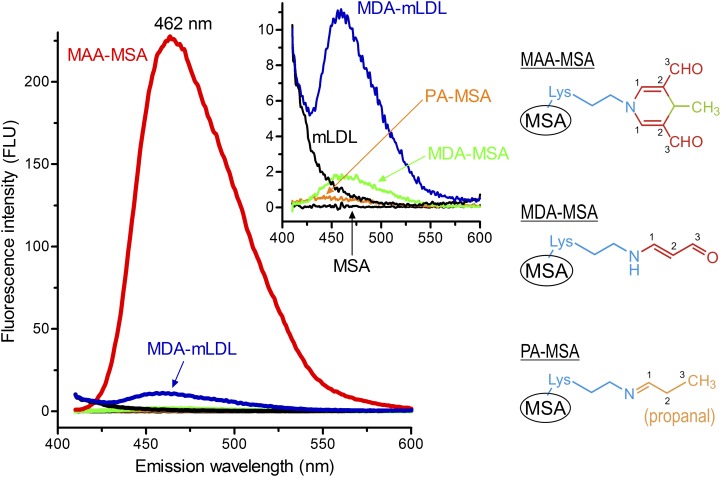
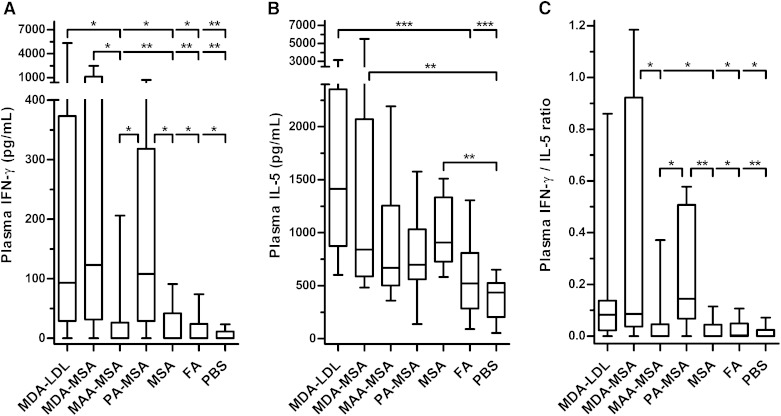
Immunization with homologous malondialdehyde (MDA)-modified LDL (MDA-LDL) leads to atheroprotection in experimental models supporting the concept that a vaccine to oxidation-specific epitopes (OSEs) of oxidized LDL could limit atherogenesis. However, modification of human LDL with OSE to use as an immunogen would be impractical for generalized use. Furthermore, when MDA is used to modify LDL, a wide variety of related MDA adducts are formed, both simple and more complex. To define the relevant epitopes that would reproduce the atheroprotective effects of immunization with MDA-LDL, we sought to determine the responsible immunodominant and atheroprotective adducts. We now demonstrate that fluorescent adducts of MDA involving the condensation of two or more MDA molecules with lysine to form malondialdehyde-acetaldehyde (MAA)-type adducts generate immunodominant epitopes that lead to atheroprotective responses. We further demonstrate that a T helper (Th) 2-biased hapten-specific humoral and cellular response is sufficient, and thus, MAA-modified homologous albumin is an equally effective immunogen. We further show that such Th2-biased humoral responses per se are not atheroprotective if they do not target relevant antigens. These data demonstrate the feasibility of development of a small-molecule immunogen that could stimulate MAA-specific immune responses, which could be used to develop a vaccine approach to retard or prevent atherogenesis.
Keywords: adaptive immunity; humoral immunity; malondialdehyde-acetaldehyde adducts; oxidation-specific epitopes.
Copyright © 2014 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures








References
-
- Glass C. K., Witztum J. L. 2001. Atherosclerosis. The road ahead. Cell. 104: 503–516. - PubMed
-
- Libby P., Ridker P. M., Hansson G. K. 2011. Progress and challenges in translating the biology of atherosclerosis. Nature. 473: 317–325. - PubMed
-
- Miller Y. I., Choi S. H., Wiesner P., Fang L., Harkewicz R., Hartvigsen K., Boullier A., Gonen A., Diehl C. J., Que X., et al. 2011. Oxidation-specific epitopes are danger-associated molecular patterns recognized by pattern recognition receptors of innate immunity. Circ. Res. 108: 235–248. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
