

Primary cilia signaling mediates intraocular pressure sensation
- PMID: 25143588
- PMCID: PMC4156748
- DOI: 10.1073/pnas.1323292111
Primary cilia signaling mediates intraocular pressure sensation
Abstract
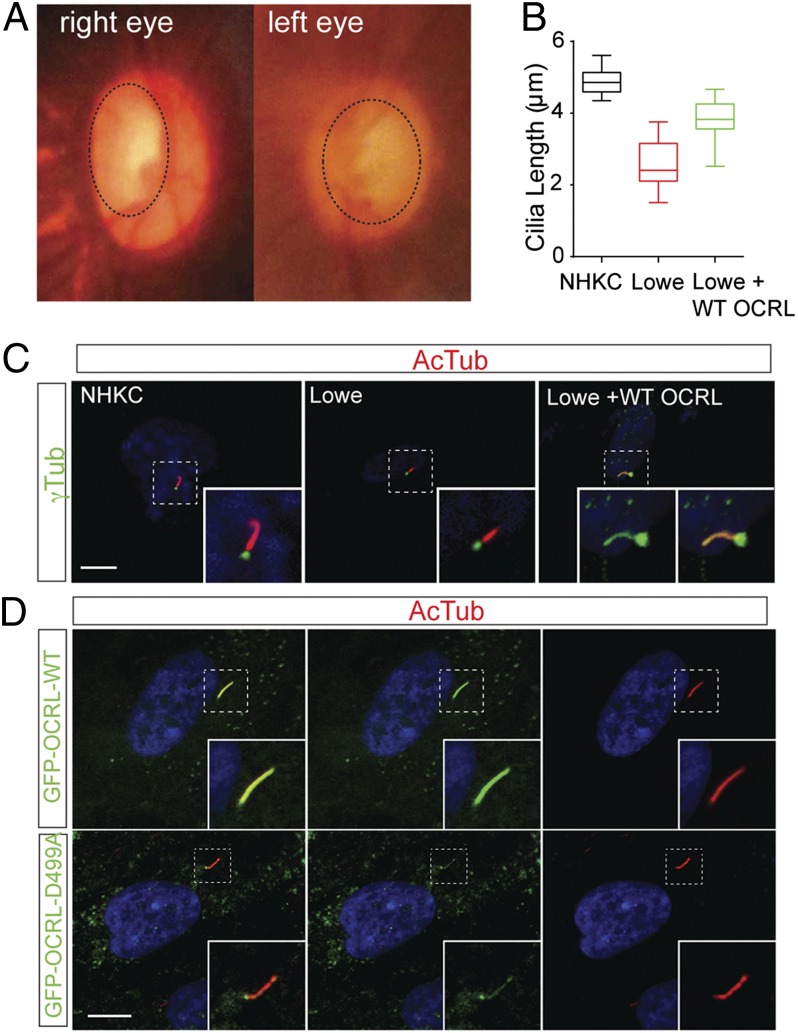
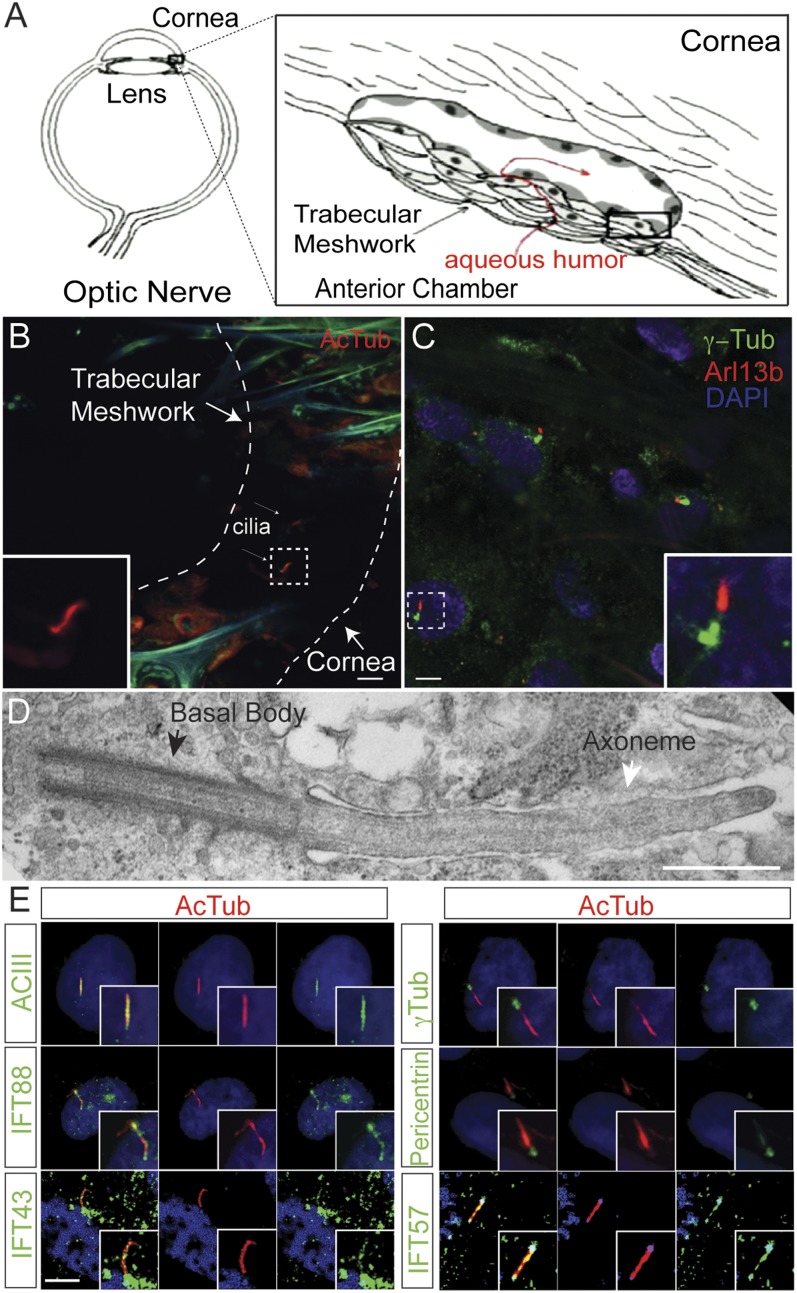
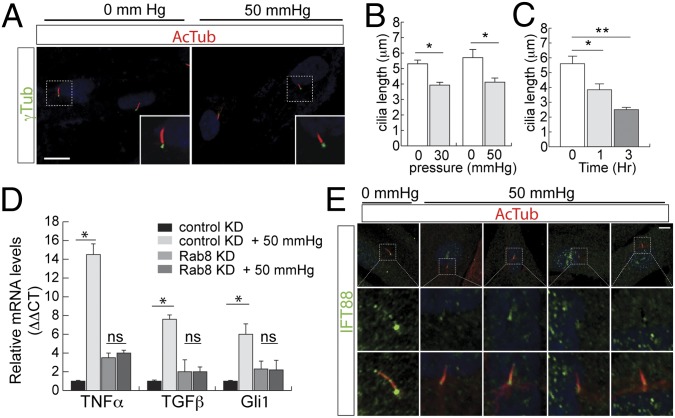
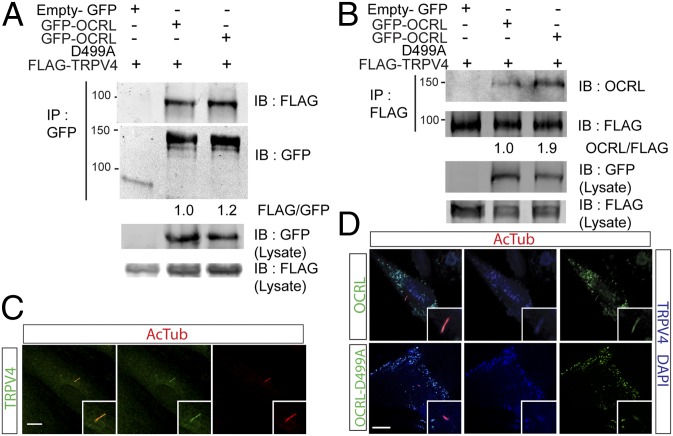
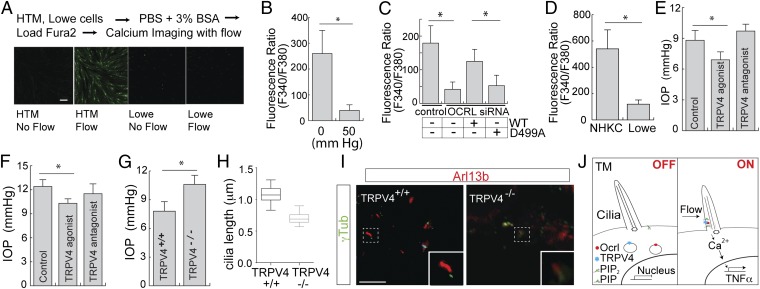
Lowe syndrome is a rare X-linked congenital disease that presents with congenital cataracts and glaucoma, as well as renal and cerebral dysfunction. OCRL, an inositol polyphosphate 5-phosphatase, is mutated in Lowe syndrome. We previously showed that OCRL is involved in vesicular trafficking to the primary cilium. Primary cilia are sensory organelles on the surface of eukaryotic cells that mediate mechanotransduction in the kidney, brain, and bone. However, their potential role in the trabecular meshwork (TM) in the eye, which regulates intraocular pressure, is unknown. Here, we show that TM cells, which are defective in glaucoma, have primary cilia that are critical for response to pressure changes. Primary cilia in TM cells shorten in response to fluid flow and elevated hydrostatic pressure, and promote increased transcription of TNF-α, TGF-β, and GLI1 genes. Furthermore, OCRL is found to be required for primary cilia to respond to pressure stimulation. The interaction of OCRL with transient receptor potential vanilloid 4 (TRPV4), a ciliary mechanosensory channel, suggests that OCRL may act through regulation of this channel. A novel disease-causing OCRL allele prevents TRPV4-mediated calcium signaling. In addition, TRPV4 agonist GSK 1016790A treatment reduced intraocular pressure in mice; TRPV4 knockout animals exhibited elevated intraocular pressure and shortened cilia. Thus, mechanotransduction by primary cilia in TM cells is implicated in how the eye senses pressure changes and highlights OCRL and TRPV4 as attractive therapeutic targets for the treatment of glaucoma. Implications of OCRL and TRPV4 in primary cilia function may also shed light on mechanosensation in other organ systems.
Conflict of interest statement
Conflict of interest statement: Y.S. is an inventor on a patent related to the work described, which is managed by Indiana University School of Medicine.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- NIH/NHL115140/PHS HHS/United States
- NIH/NCI151765/PHS HHS/United States
- NIH/NEI22639/PHS HHS/United States
- R01 CA151765/CA/NCI NIH HHS/United States
- R01 HL115140/HL/NHLBI NIH HHS/United States
- R01 ES020866/ES/NIEHS NIH HHS/United States
- NHL P01-056693/PHS HHS/United States
- P30 EY008126/EY/NEI NIH HHS/United States
- 5P30EY08126/EY/NEI NIH HHS/United States
- R01 EY022639/EY/NEI NIH HHS/United States
- KL2 TR001106/TR/NCATS NIH HHS/United States
- R01 EY025295/EY/NEI NIH HHS/United States
- NIH/NEI022058/PHS HHS/United States
- K08 EY022058/EY/NEI NIH HHS/United States
- NIH/KL2TR001106/TR/NCATS NIH HHS/United States
- UL1 TR001108/TR/NCATS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
