

MondoA deficiency enhances sprint performance in mice
- PMID: 25145386
- PMCID: PMC4410994
- DOI: 10.1042/BJ20140530
MondoA deficiency enhances sprint performance in mice
Abstract
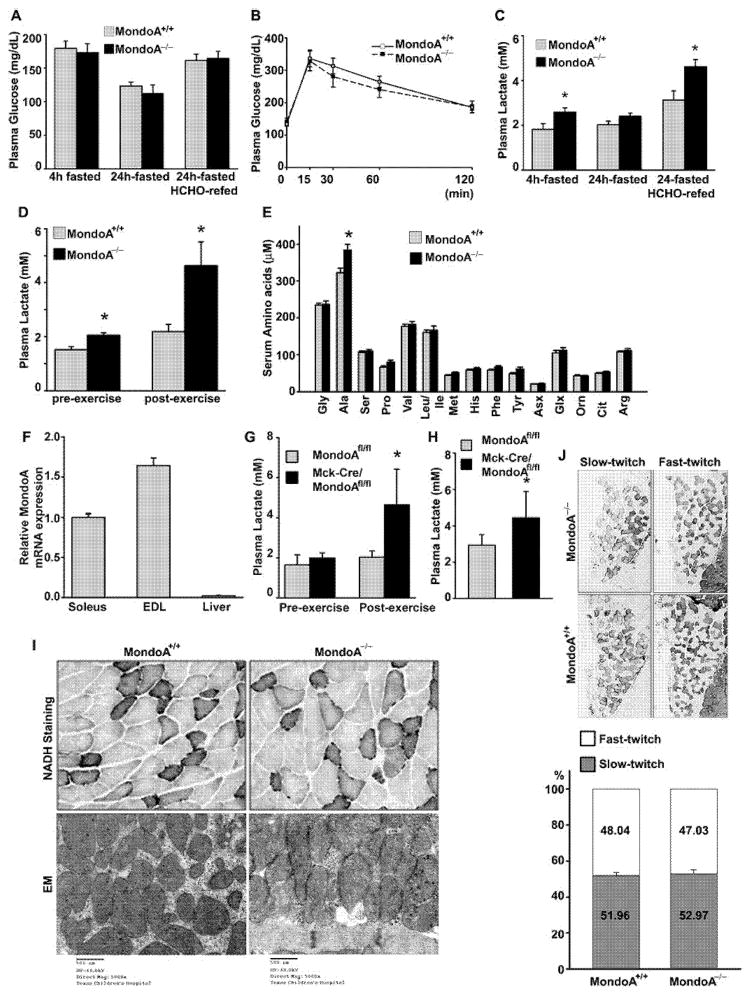
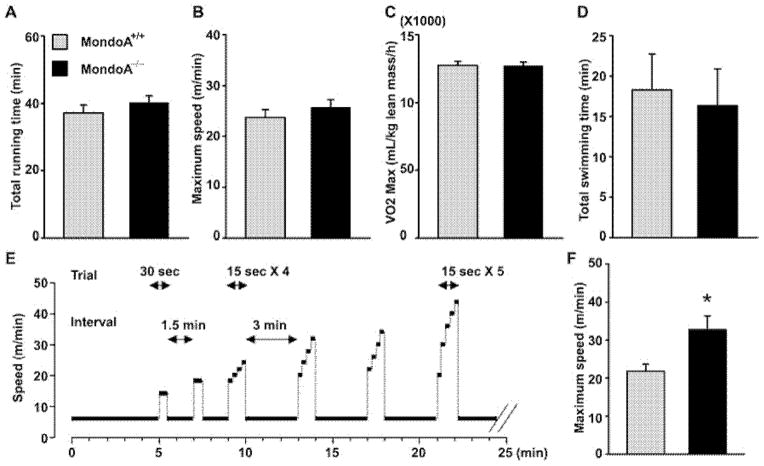
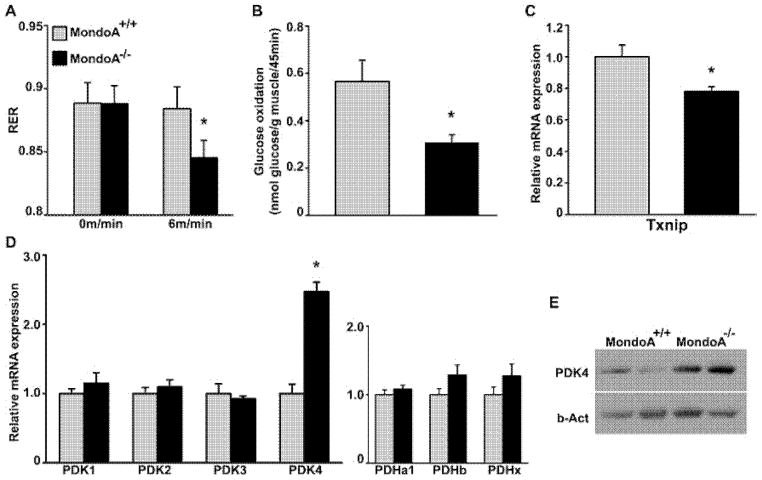
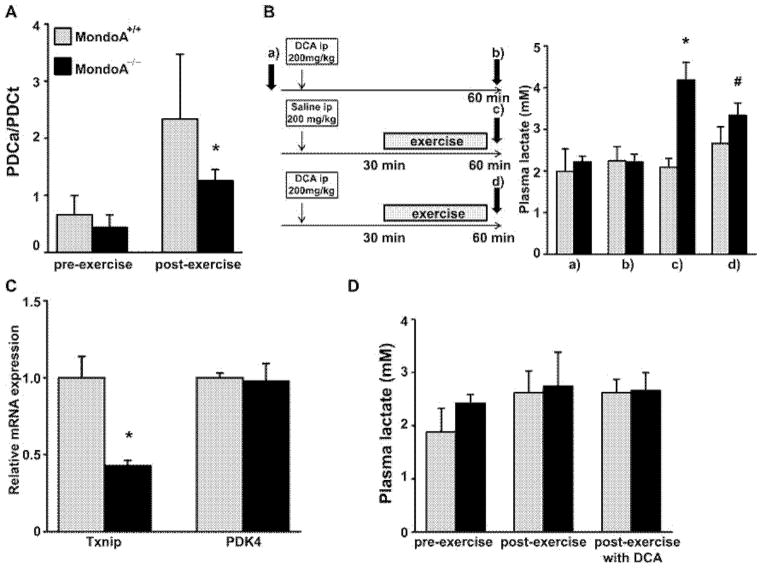
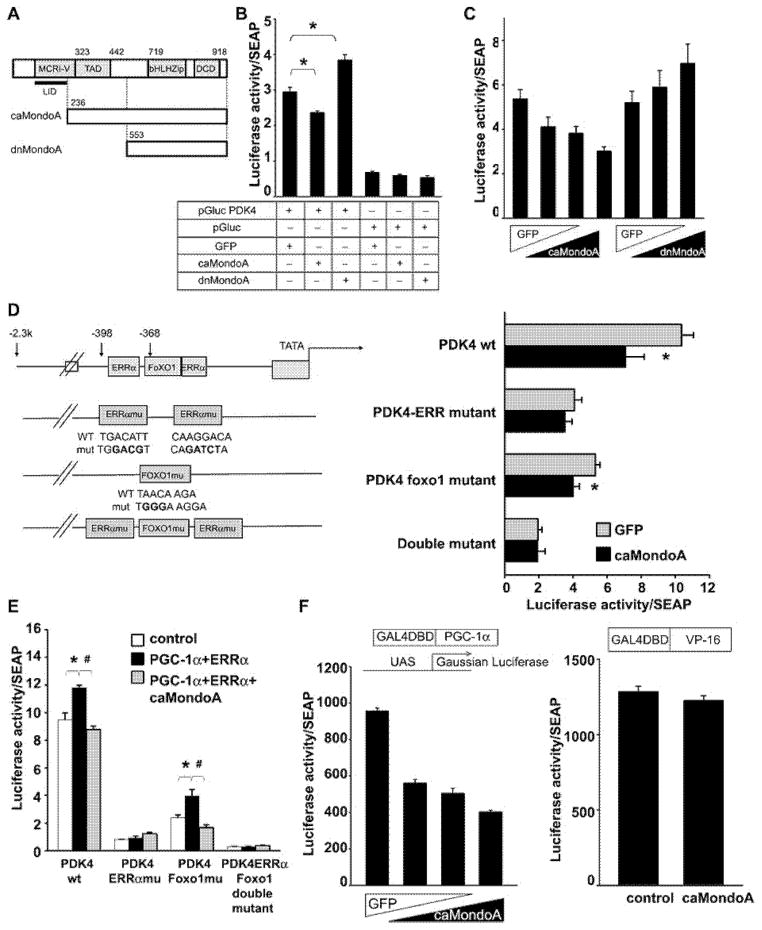
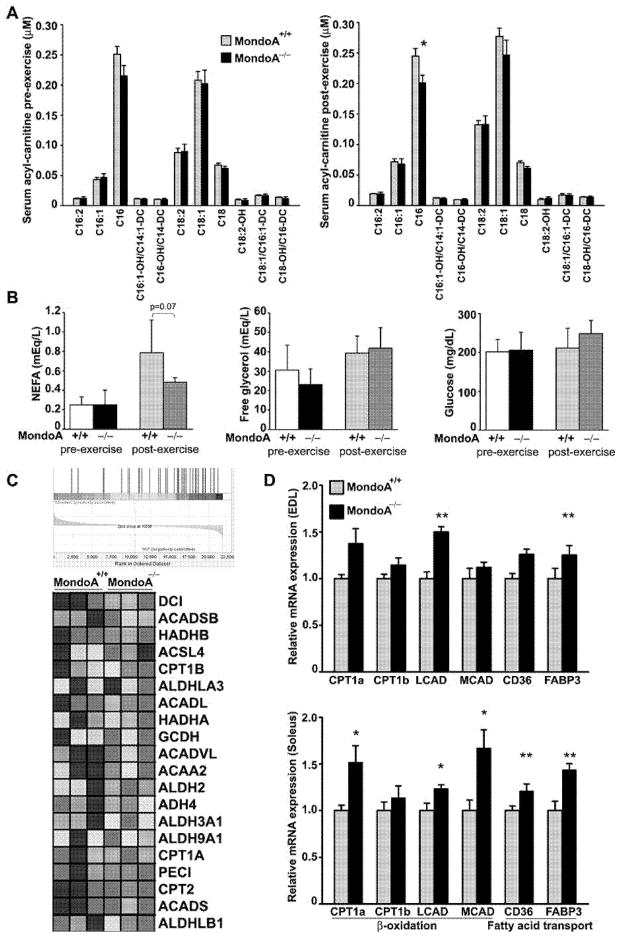
MondoA is a basic helix-loop-helix (bHLH)/leucine zipper (ZIP) transcription factor that is expressed predominantly in skeletal muscle. Studies in vitro suggest that the Max-like protein X (MondoA:Mlx) heterodimer senses the intracellular energy status and directly targets the promoter region of thioredoxin interacting protein (Txnip) and possibly glycolytic enzymes. We generated MondoA-inactivated (MondoA-/-) mice by gene targeting. MondoA-/- mice had normal body weight at birth, exhibited normal growth and appeared to be healthy. However, they exhibited unique metabolic characteristics. MondoA-/- mice built up serum lactate and alanine levels and utilized fatty acids for fuel during exercise. Gene expression and promoter analysis suggested that MondoA functionally represses peroxisome-proliferator-activated receptor γ co-activator-1α (PGC-1α)-mediated activation of pyruvate dehydrogenase kinase 4 (PDK-4) transcription. PDK4 normally down-regulates the activity of pyruvate dehydrogenase, an enzyme complex that catalyses the decarboxylation of pyruvate to acetyl-CoA for entry into the Krebs cycle; in the absence of MondoA, pyruvate is diverted towards lactate and alanine, both products of glycolysis. Dynamic testing revealed that MondoA-/- mice excel in sprinting as their skeletal muscles display an enhanced glycolytic capacity. Our studies uncover a hitherto unappreciated function of MondoA in fuel selection in vivo. Lack of MondoA results in enhanced exercise capacity with sprinting.
Figures
References
-
- Girard J, Ferré P, Foufelle F. Mechanisms by which carbohydrates regulate expression of genes for glycolytic and lipogenic enzymes. Annu Rev Nutr. 1997;17:325–352. - PubMed
-
- Uyeda K, Yamashita H, Kawaguchi T. Carbohydrate responsive element-binding protein (ChREBP): a key regulator of glucose metabolism and fat storage. Biochem Pharmacol. 2002;63:2075–2080. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases