

Robust quantification of orientation selectivity and direction selectivity
- PMID: 25147504
- PMCID: PMC4123790
- DOI: 10.3389/fncir.2014.00092
Robust quantification of orientation selectivity and direction selectivity
Abstract
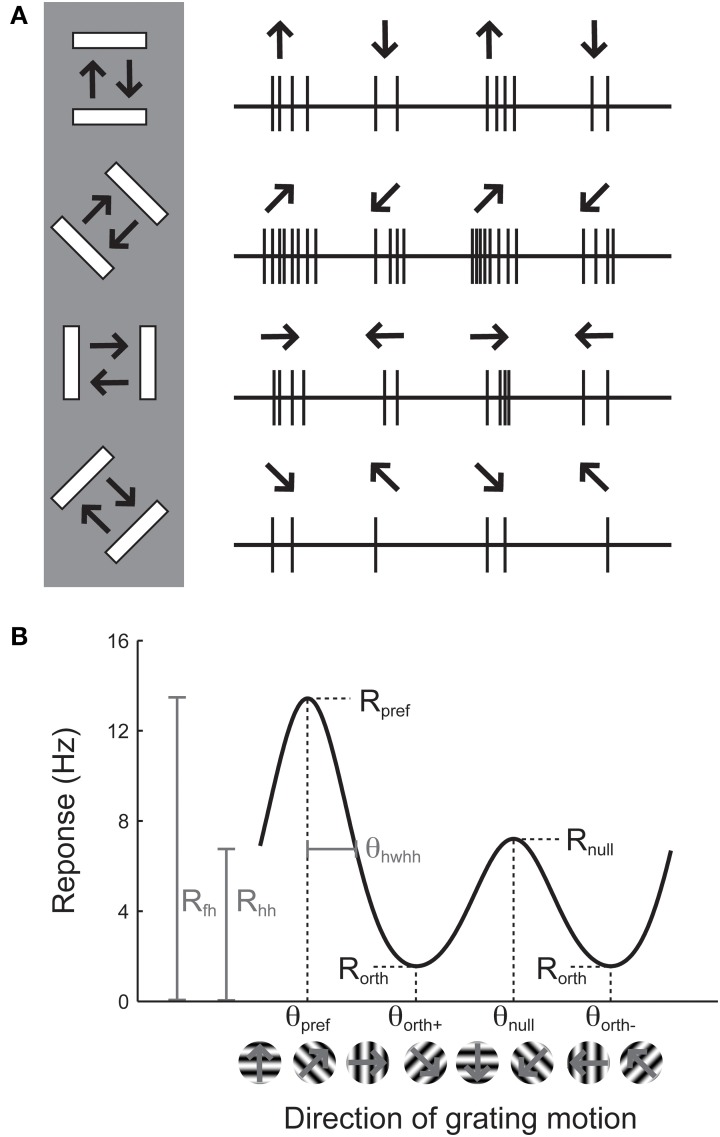
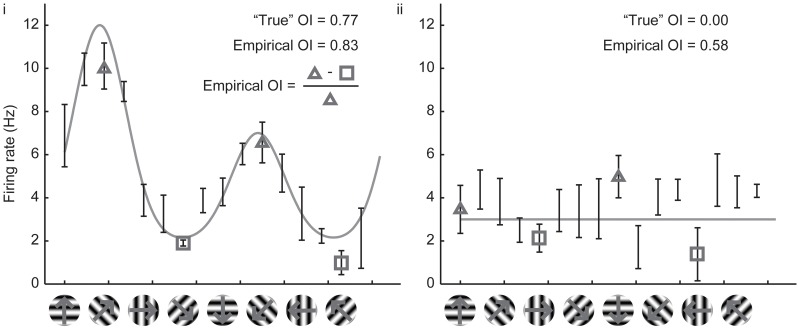
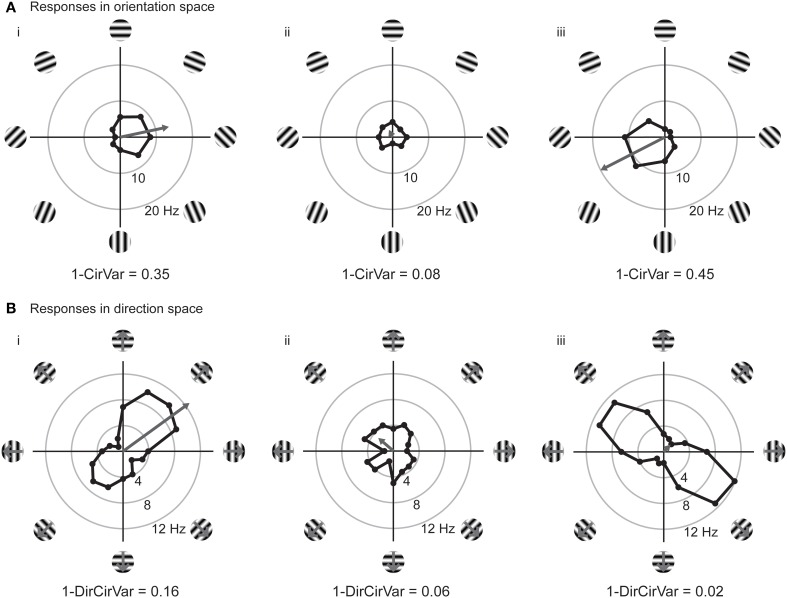
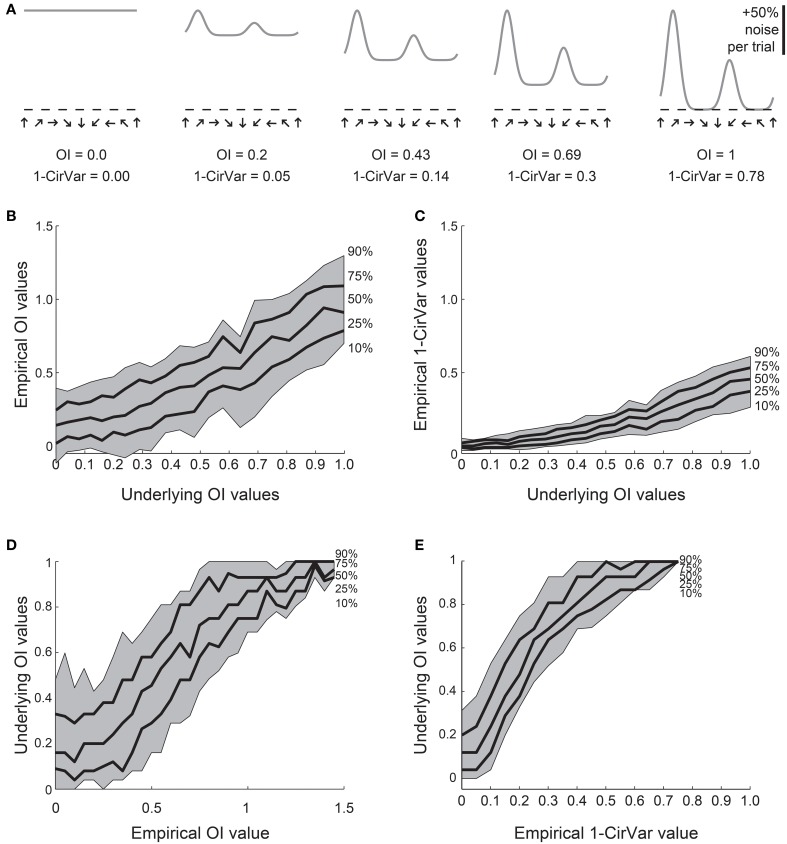
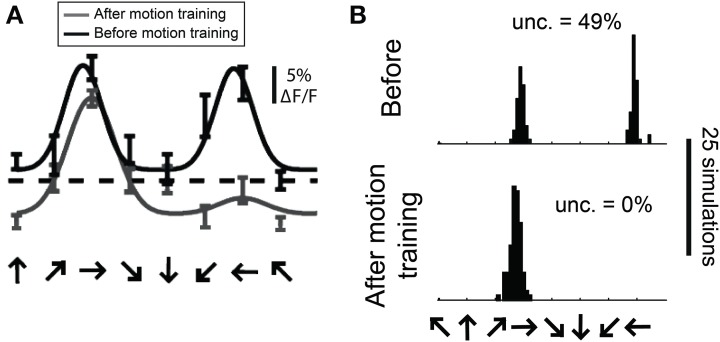
Neurons in the visual cortex of all examined mammals exhibit orientation or direction tuning. New imaging techniques are allowing the circuit mechanisms underlying orientation and direction selectivity to be studied with clarity that was not possible a decade ago. However, these new techniques bring new challenges: robust quantitative measurements are needed to evaluate the findings from these studies, which can involve thousands of cells of varying response strength. Here we show that traditional measures of selectivity such as the orientation index (OI) and direction index (DI) are poorly suited for quantitative evaluation of orientation and direction tuning. We explore several alternative methods for quantifying tuning and for addressing a variety of questions that arise in studies on orientation- and direction-tuned cells and cell populations. We provide recommendations for which methods are best suited to which applications and we offer tips for avoiding potential pitfalls in applying these methods. Our goal is to supply a solid quantitative foundation for studies involving orientation and direction tuning.
Keywords: Monte Carlo; neural data analysis; sampling.
Figures
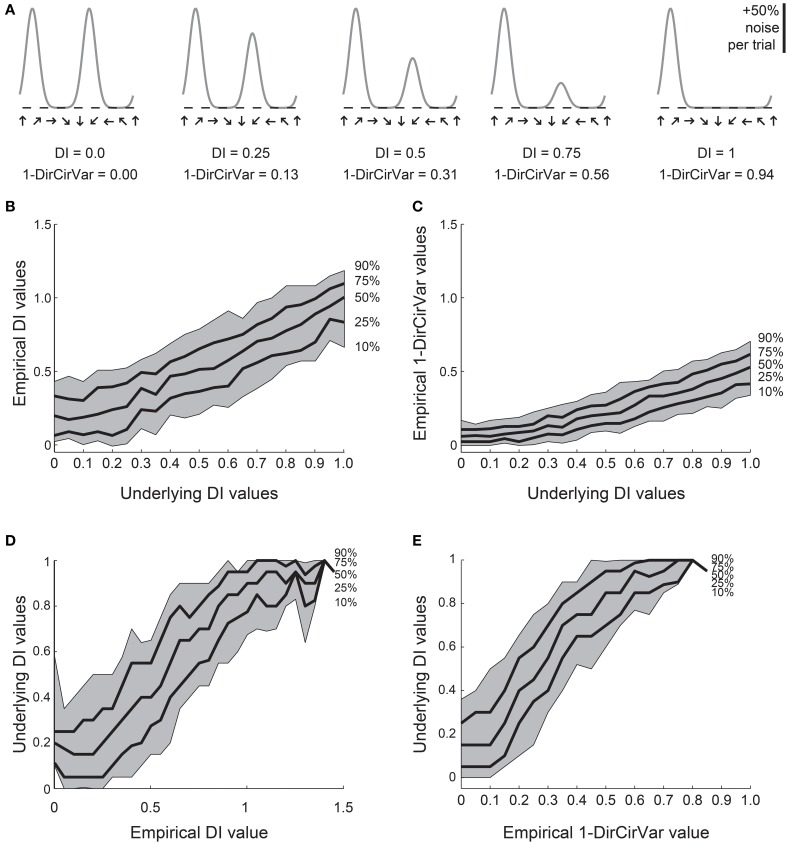
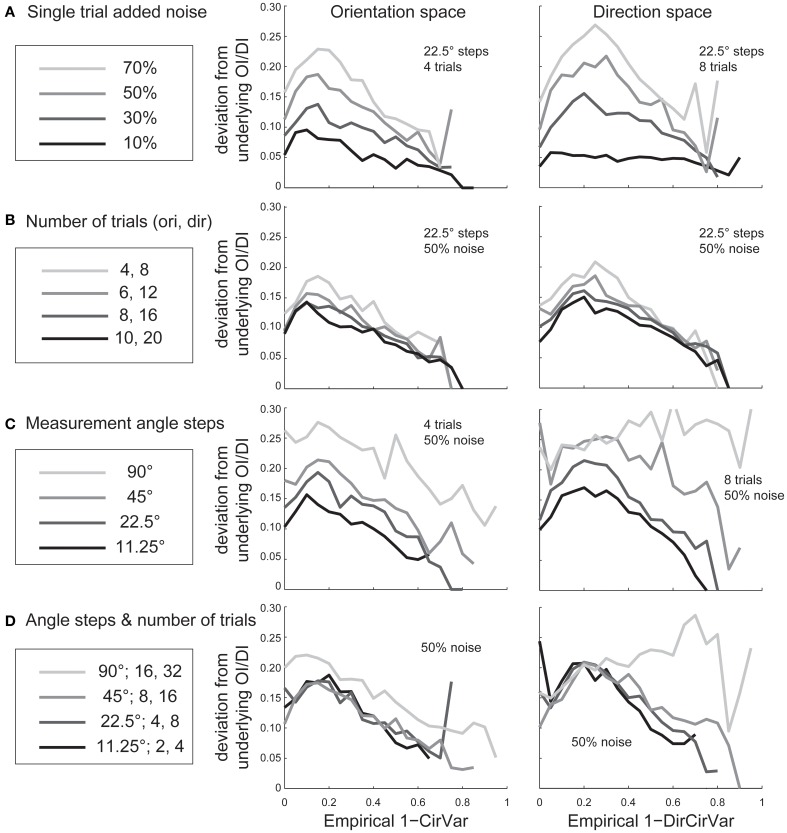
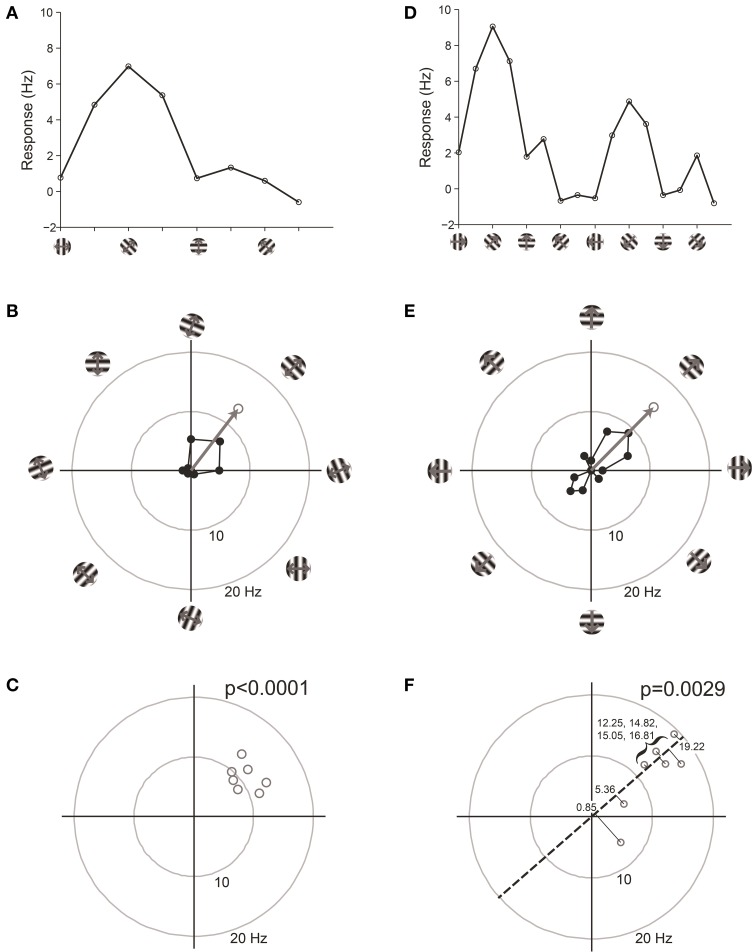
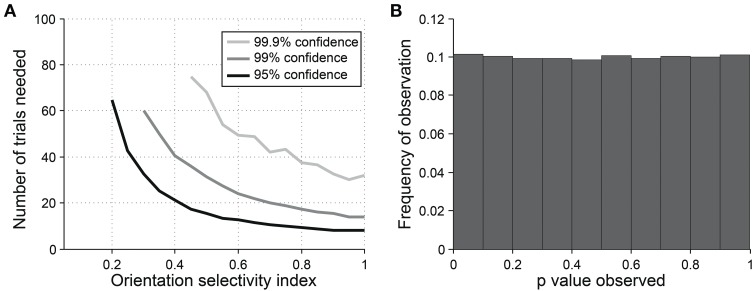
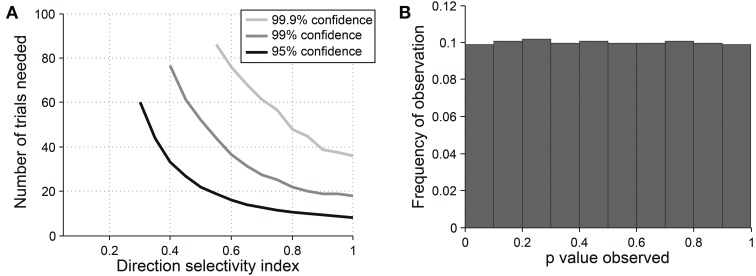
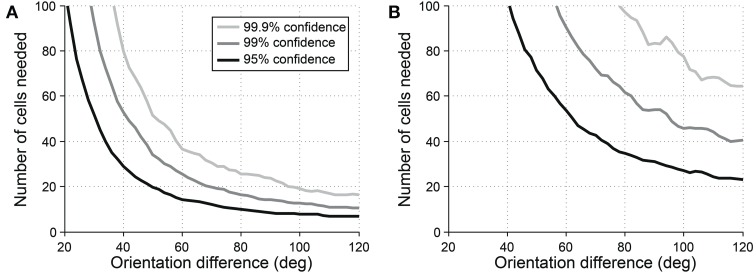
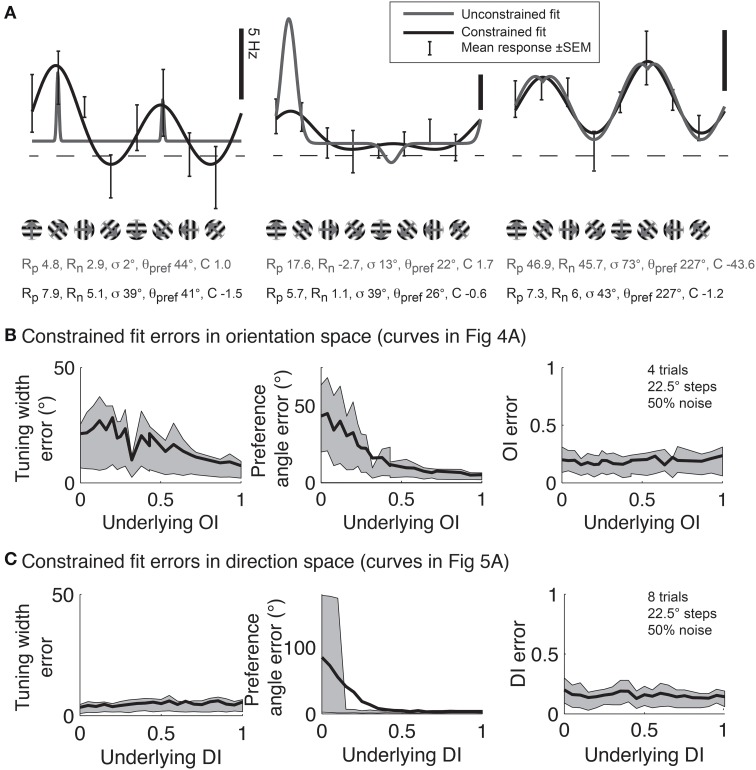
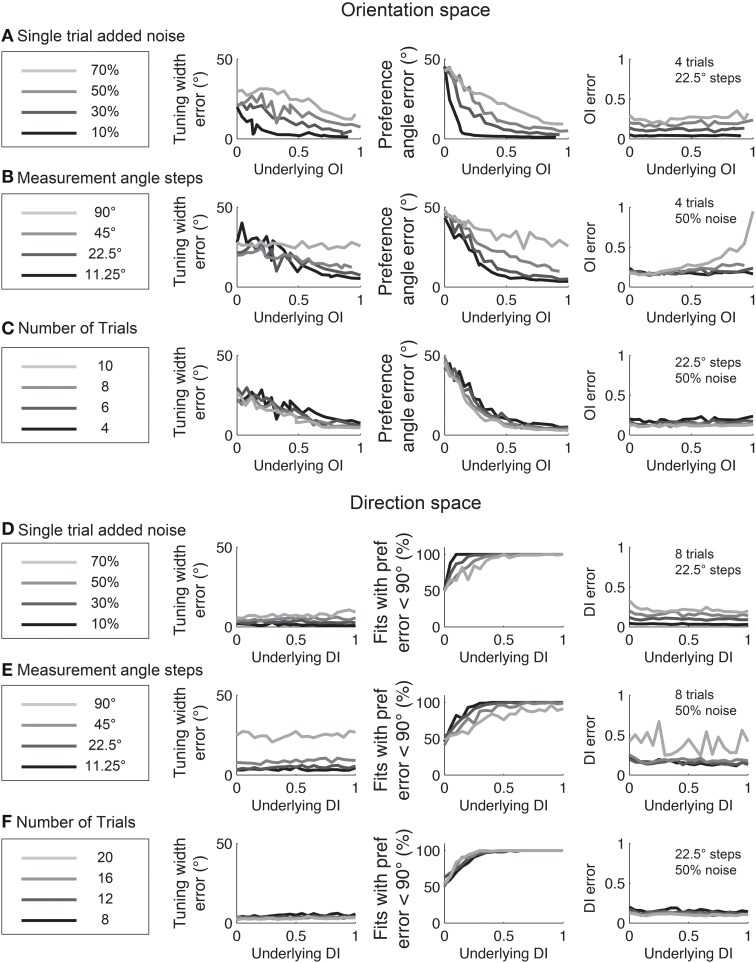
Comment in
-
Commentary: Robust quantification of orientation selectivity and direction selectivity.Front Neural Circuits. 2016 Mar 31;10:25. doi: 10.3389/fncir.2016.00025. eCollection 2016. Front Neural Circuits. 2016. PMID: 27065814 Free PMC article. No abstract available.
References
-
- Batschelet E. (1981). Circular Statistics in Biology. New York, NY: Academic Press
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources