

The kinase p38 activated by the metabolic regulator AMPK and scaffold TAB1 drives the senescence of human T cells
- PMID: 25151490
- PMCID: PMC4190666
- DOI: 10.1038/ni.2981
The kinase p38 activated by the metabolic regulator AMPK and scaffold TAB1 drives the senescence of human T cells
Abstract
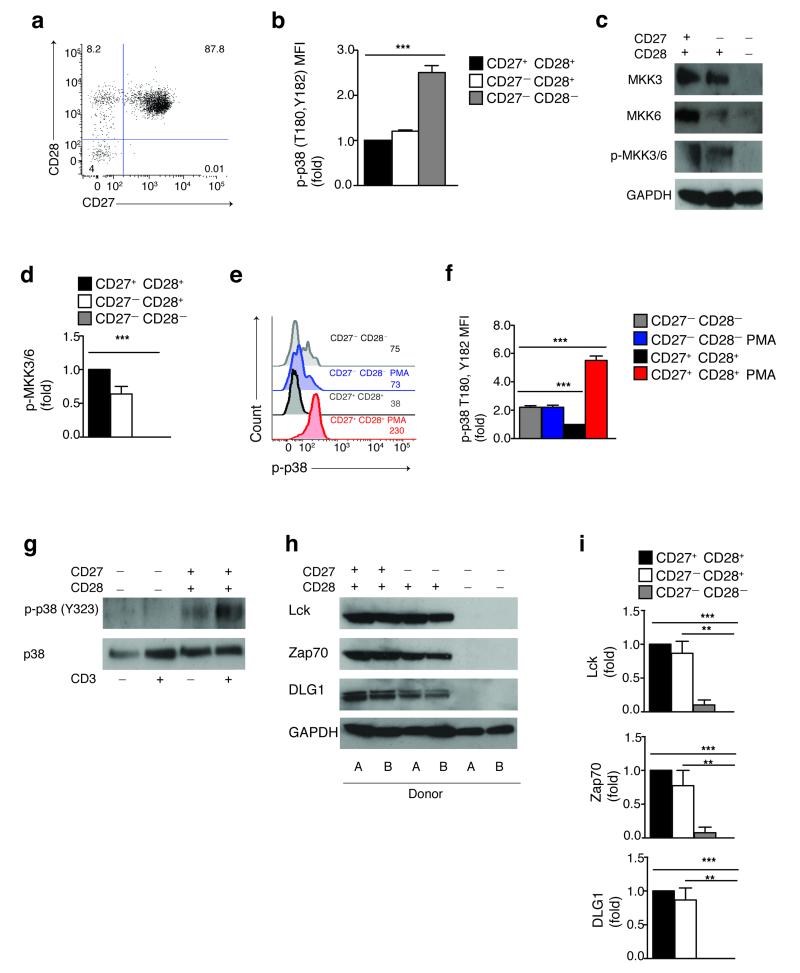
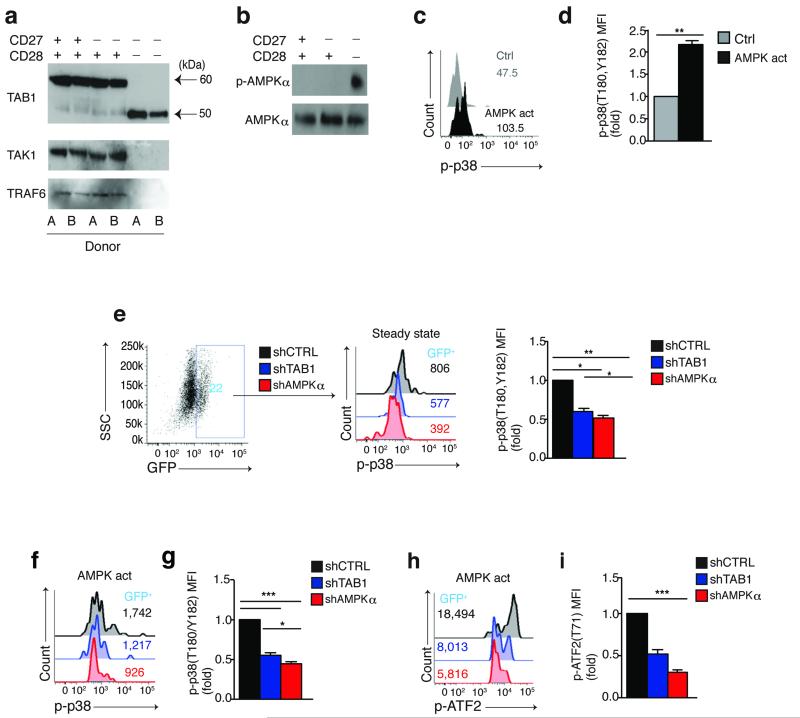
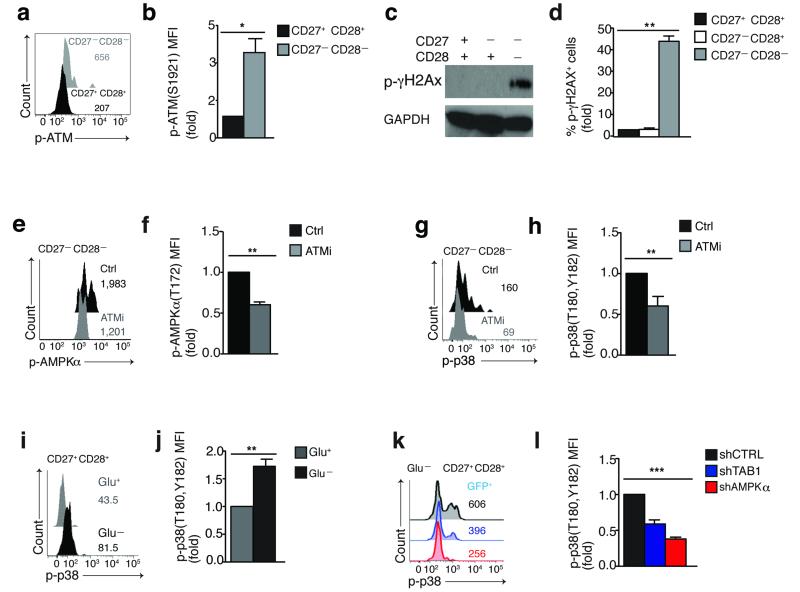
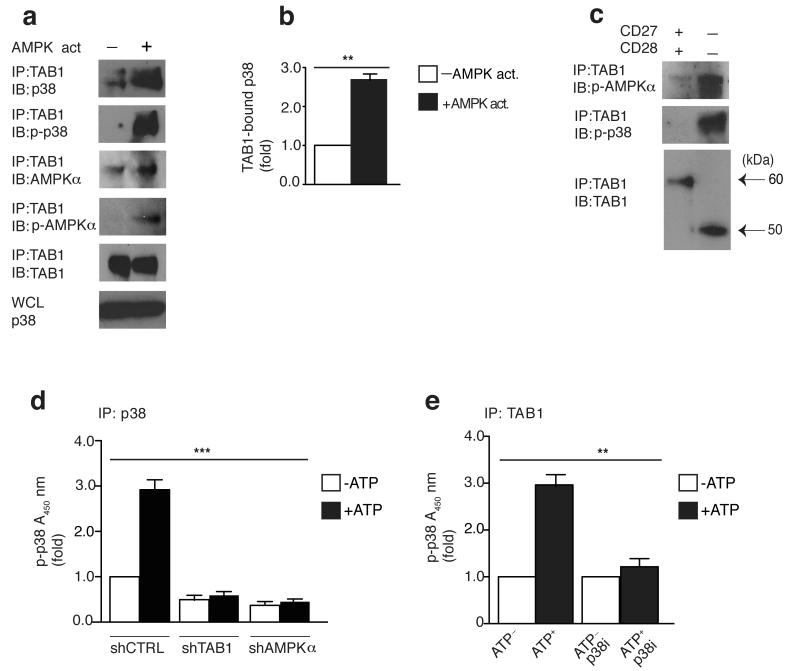
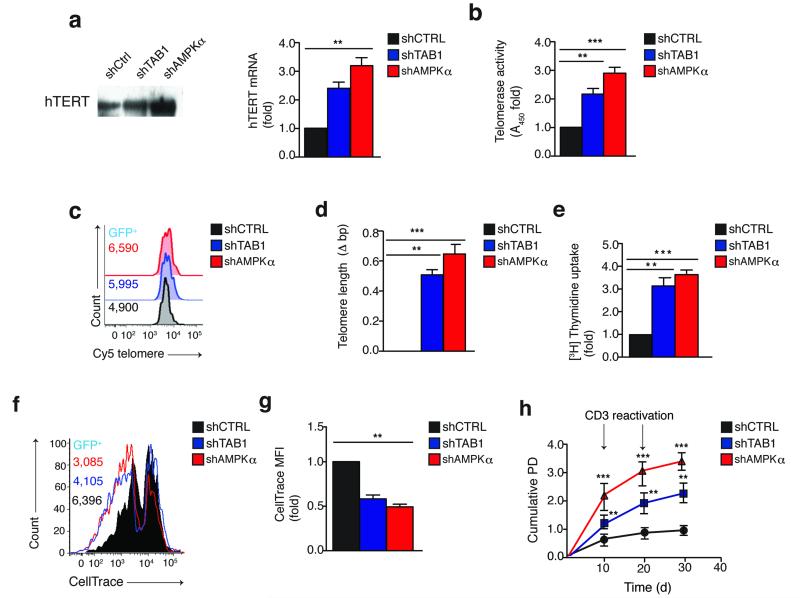
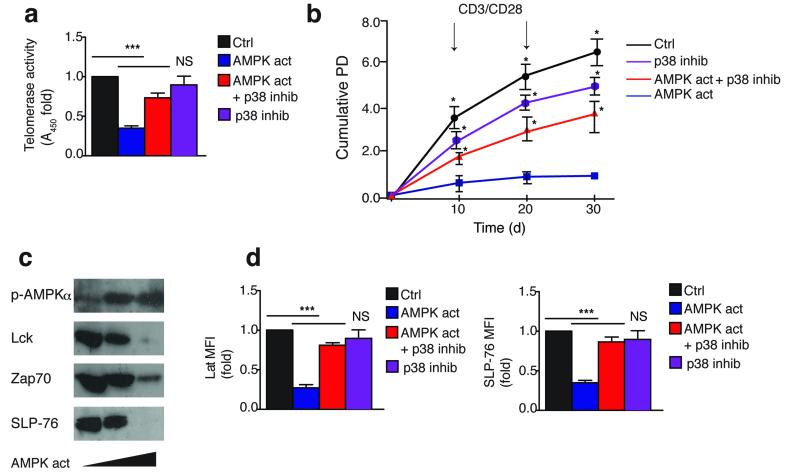
In T lymphocytes, the mitogen-activated protein kinase (MAPK) p38 regulates pleiotropic functions and is activated by canonical MAPK signaling or the alternative activation pathway downstream of the T cell antigen receptor (TCR). Here we found that senescent human T cells lacked the canonical and alternative pathways for the activation of p38 but spontaneously engaged the metabolic master regulator AMPK to trigger recruitment of p38 to the scaffold protein TAB1, which caused autophosphorylation of p38. Signaling via this pathway inhibited telomerase activity, T cell proliferation and the expression of key components of the TCR signalosome. Our findings identify a previously unrecognized mode for the activation of p38 in T cells driven by intracellular changes such as low-nutrient and DNA-damage signaling (an 'intrasensory' pathway). The proliferative defect of senescent T cells was reversed by blockade of AMPK-TAB1-dependent activation of p38.
Figures
References
-
- Huang Y, Wange RL. T cell receptor signaling: beyond complex complexes. The Journal of biological chemistry. 2004;279:28827–28830. - PubMed
-
- Akbar AN, Beverley PC, Salmon M. Will telomere erosion lead to a loss of T-cell memory? Nature reviews. Immunology. 2004;4:737–743. - PubMed
-
- Fletcher JM, et al. Cytomegalovirus-specific CD4+ T cells in healthy carriers are continuously driven to replicative exhaustion. J Immunol. 2005;175:8218–8225. - PubMed
-
- Henson SM, et al. KLRG1 signaling induces defective Akt (ser473) phosphorylation and proliferative dysfunction of highly differentiated CD8+ T cells. Blood. 2009;113:6619–6628. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
