

Histone deacetylase 11: A novel epigenetic regulator of myeloid derived suppressor cell expansion and function
- PMID: 25155994
- PMCID: PMC4252813
- DOI: 10.1016/j.molimm.2014.08.002
Histone deacetylase 11: A novel epigenetic regulator of myeloid derived suppressor cell expansion and function
Abstract
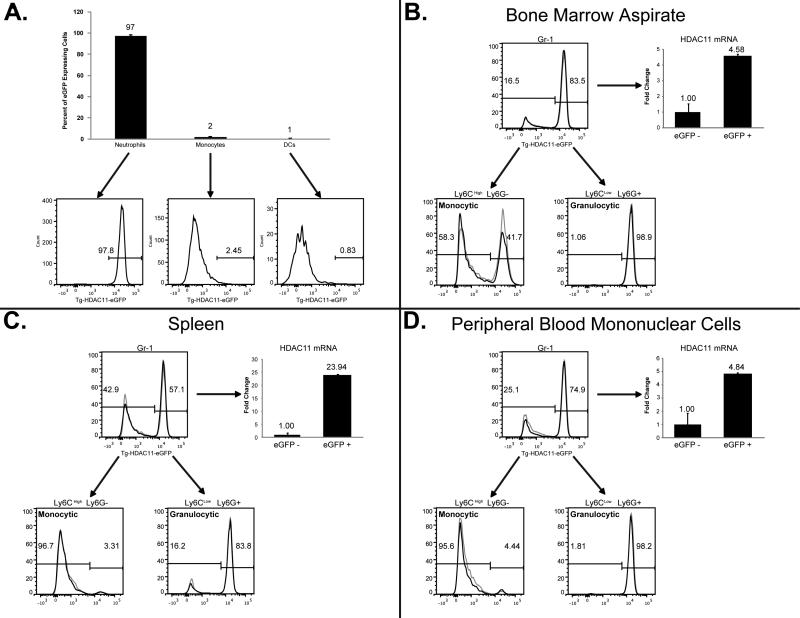
Myeloid-derived suppressor cells (MDSCs), a heterogeneous population of cells capable of suppressing anti-tumor T cell function in the tumor microenvironment, represent an imposing obstacle in the development of cancer immunotherapeutics. Thus, identifying elements essential to the development and perpetuation of these cells will undoubtedly improve our ability to circumvent their suppressive impact. HDAC11 has emerged as a key regulator of IL-10 gene expression in myeloid cells, suggesting that this may represent an important targetable axis through which to dampen MDSC formation. Using a murine transgenic reporter model system where eGFP expression is controlled by the HDAC11 promoter (Tg-HDAC11-eGFP), we provide evidence that HDAC11 appears to function as a negative regulator of MDSC expansion/function in vivo. MDSCs isolated from EL4 tumor-bearing Tg-HDAC11-eGFP display high expression of eGFP, indicative of HDAC11 transcriptional activation at steady state. In striking contrast, immature myeloid cells in tumor-bearing mice display a diminished eGFP expression, implying that the transition of IMC to MDSC's require a decrease in the expression of HDAC11, where we postulate that it acts as a gate-keeper of myeloid differentiation. Indeed, tumor-bearing HDAC11-knockout mice (HDAC11-KO) demonstrate a more suppressive MDSC population as compared to wild-type (WT) tumor-bearing control. Notably, the HDAC11-KO tumor-bearing mice exhibit enhanced tumor growth kinetics when compare to the WT control mice. Thus, through a better understanding of this previously unknown role of HDAC11 in MDSC expansion and function, rational development of targeted epigenetic modifiers may allow us to thwart a powerful barrier to efficacious immunotherapies.
Keywords: HDAC11; Immuno-suppression; MDSCs; Myelopoiesis.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures





References
-
- Almand B, Clark JI, Nikitina E, van Beynen J, English NR, Knight SC, Carbone DP, Gabrilovich DI. Increased production of immature myeloid cells in cancer patients: a mechanism of immunosuppression in cancer. J Immunol. 2001;166(1):678–689. - PubMed
-
- Basu S, Hodgson G, Zhang HH, Katz M, Quilici C, Dunn AR. Emergency” granulopoiesis in G-CSF-deficient mice in response to Candida albicans infection. Blood. 2000;95(12):3725–3733. - PubMed
-
- Bronte V, Zanovello P. Regulation of immune responses by L-arginine metabolism. Nat Rev Immunol. 2005;5(8):641–654. - PubMed
-
- Cheng F, Lienlaf M, Perez-Villarroel P, Wang HW, Lee C, Woan K, Woods D, Knox T, Bergman J, Pinilla-Ibarz J, Kozikowski A, Seto E, Sotomayor EM, Villagra A. Divergent roles of histone deacetylase 6 (HDAC6) and histone deacetylase 11 (HDAC11) on the transcriptional regulation of IL10 in antigen presenting cells. Mol Immunol. 2014;60(1):44–53. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
