

K⁺ channel gating: C-type inactivation is enhanced by calcium or lanthanum outside
- PMID: 25156116
- PMCID: PMC4144669
- DOI: 10.1085/jgp.201411223
K⁺ channel gating: C-type inactivation is enhanced by calcium or lanthanum outside
Abstract
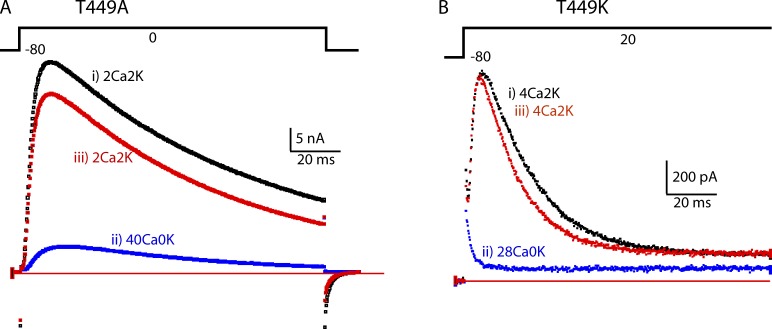
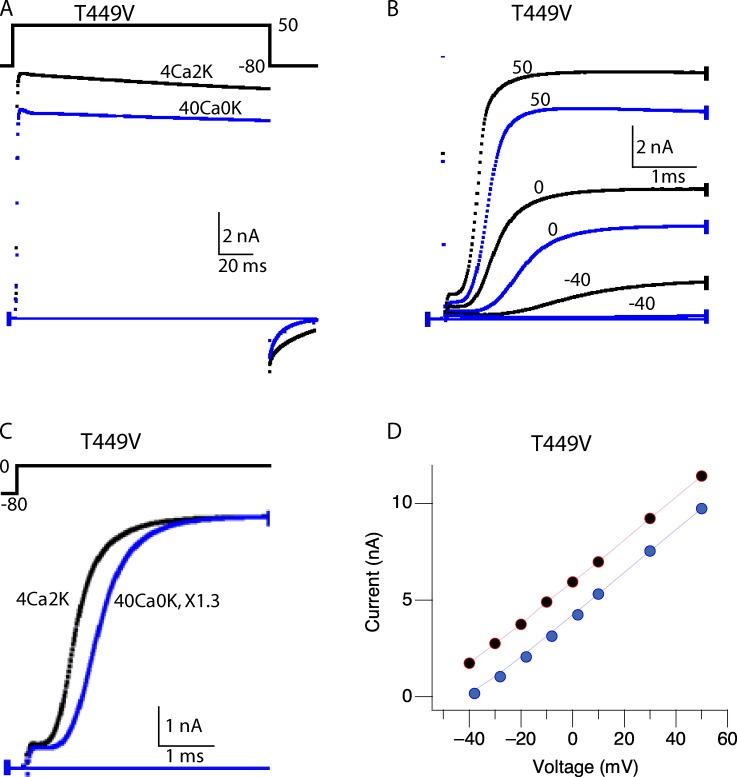
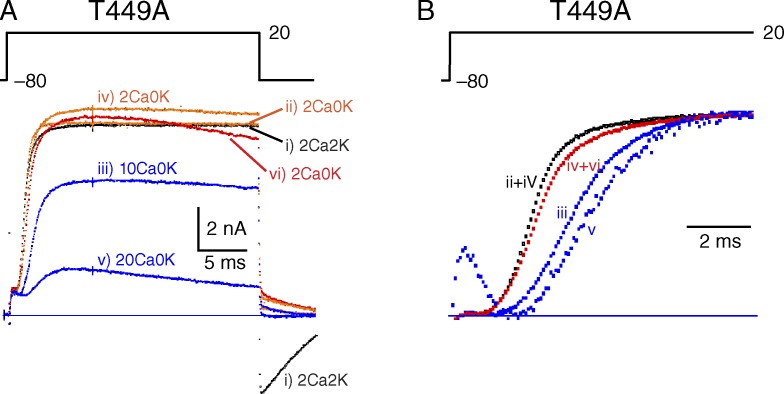
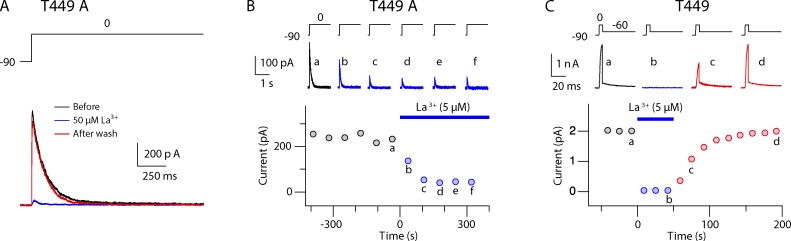
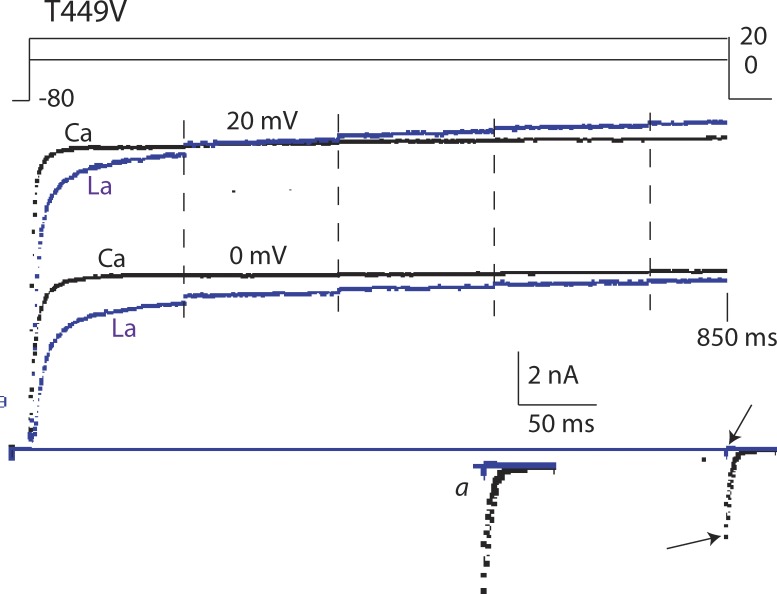
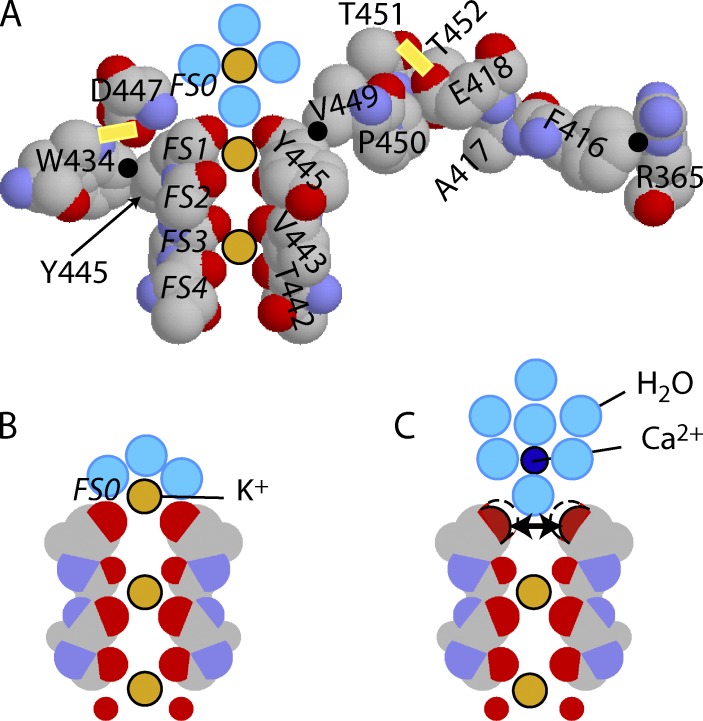
Many voltage-gated K(+) channels exhibit C-type inactivation. This typically slow process has been hypothesized to result from dilation of the outer-most ring of the carbonyls in the selectivity filter, destroying this ring's ability to bind K(+) with high affinity. We report here strong enhancement of C-type inactivation upon extracellular addition of 10-40 mM Ca(2+) or 5-50 µM La(3+). These multivalent cations mildly increase the rate of C-type inactivation during depolarization and markedly promote inactivation and/or suppress recovery when membrane voltage (V(m)) is at resting levels (-80 to -100 mV). At -80 mV with 40 mM Ca(2+) and 0 mM K(+) externally, ShBΔN channels with the mutation T449A inactivate almost completely within 2 min or less with no pulsing. This behavior is observed only in those mutants that show C-type inactivation on depolarization and is distinct from the effects of Ca(2+) and La(3+) on activation (opening and closing of the V(m)-controlled gate), i.e., slower activation of K(+) channels and a positive shift of the mid-voltage of activation. The Ca(2+)/La(3+) effects on C-type inactivation are antagonized by extracellular K(+) in the low millimolar range. This, together with the known ability of Ca(2+) and La(3+) to block inward current through K(+) channels at negative voltage, strongly suggests that Ca(2+)/La(3+) acts at the outer mouth of the selectivity filter. We propose that at -80 mV, Ca(2+) or La(3+) ions compete effectively with K(+) at the channel's outer mouth and prevent K(+) from stabilizing the filter's outer carbonyl ring.
© 2014 Armstrong and Hoshi.
Figures

References
-
- Burgess, J.1978. Metal Ions in Solution. Ellis Horwood, Chichester, NY: 481 pp
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
