

Unfolded protein response activation reduces secretion and extracellular aggregation of amyloidogenic immunoglobulin light chain
- PMID: 25157167
- PMCID: PMC4246986
- DOI: 10.1073/pnas.1406050111
Unfolded protein response activation reduces secretion and extracellular aggregation of amyloidogenic immunoglobulin light chain
Abstract
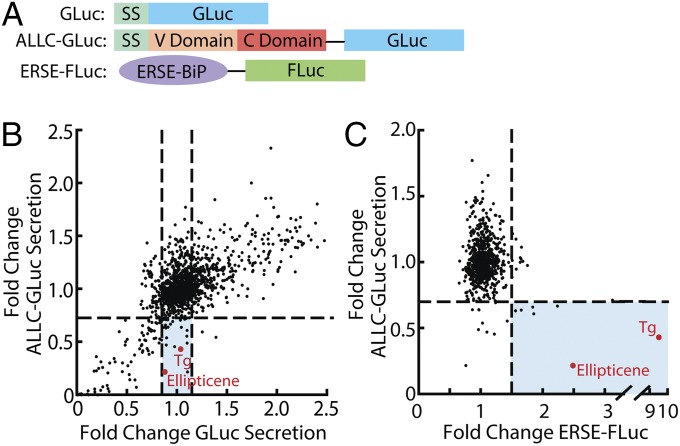
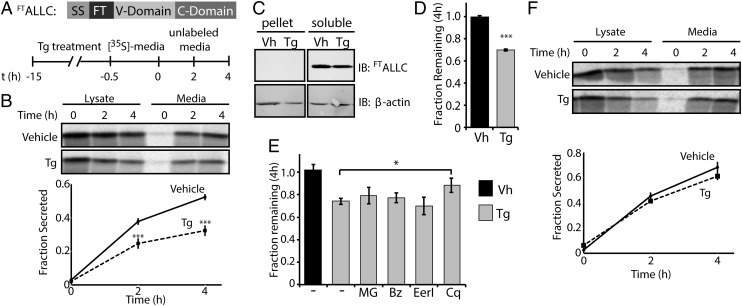
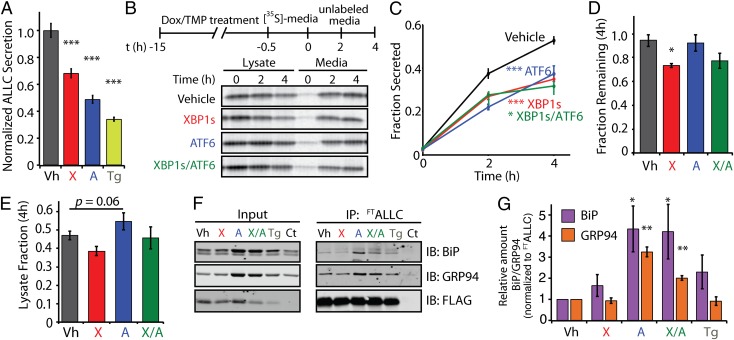
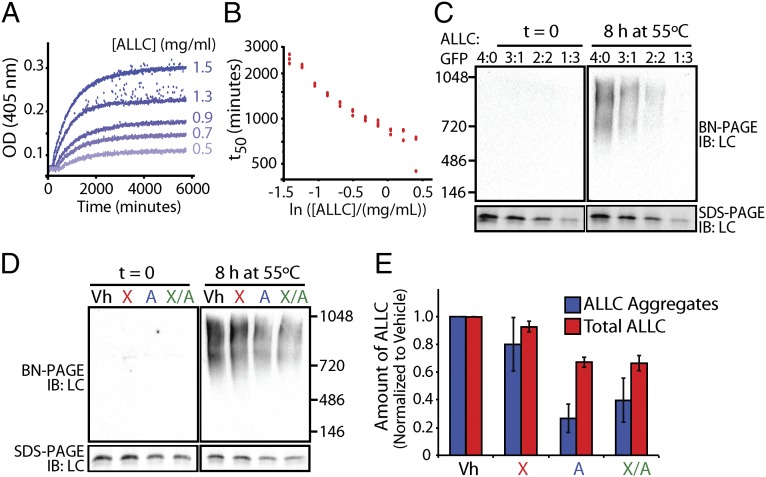
Light-chain amyloidosis (AL) is a degenerative disease characterized by the extracellular aggregation of a destabilized amyloidogenic Ig light chain (LC) secreted from a clonally expanded plasma cell. Current treatments for AL revolve around ablating the cancer plasma cell population using chemotherapy regimens. Unfortunately, this approach is limited to the ∼ 70% of patients who do not exhibit significant organ proteotoxicity and can tolerate chemotherapy. Thus, identifying new therapeutic strategies to alleviate LC organ proteotoxicity should allow AL patients with significant cardiac and/or renal involvement to subsequently tolerate established chemotherapy treatments. Using a small-molecule screening approach, the unfolded protein response (UPR) was identified as a cellular signaling pathway whose activation selectively attenuates secretion of amyloidogenic LC, while not affecting secretion of a nonamyloidogenic LC. Activation of the UPR-associated transcription factors XBP1s and/or ATF6 in the absence of stress recapitulates the selective decrease in amyloidogenic LC secretion by remodeling the endoplasmic reticulum proteostasis network. Stress-independent activation of XBP1s, or especially ATF6, also attenuates extracellular aggregation of amyloidogenic LC into soluble aggregates. Collectively, our results show that stress-independent activation of these adaptive UPR transcription factors offers a therapeutic strategy to reduce proteotoxicity associated with LC aggregation.
Keywords: ER proteostasis; amyloid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cohen AD, Comenzo RL. Systemic light-chain amyloidosis: Advances in diagnosis, prognosis, and therapy. Hematology (Am Soc Hematol Educ Program) 2010;2010:287–294. - PubMed
-
- Buxbaum JN, Linke RP. A molecular history of the amyloidoses. J Mol Biol. 2012;421(2-3):142–159. - PubMed
-
- Jones NF, Hilton PJ, Tighe JR, Hobbs JR. Treatment of “primary” renal amyloidosis with melphalan. Lancet. 1972;2(7778):616–619. - PubMed
-
- Jaccard A, et al. Myélome Autogreffe (MAG) and Intergroupe Francophone du Myélome (IFM) Intergroup High-dose melphalan versus melphalan plus dexamethasone for AL amyloidosis. N Engl J Med. 2007;357(11):1083–1093. - PubMed
-
- Kastritis E, et al. Bortezomib with or without dexamethasone in primary systemic (light chain) amyloidosis. J Clin Oncol. 2010;28(6):1031–1037. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
