

SORCS1 is necessary for normal insulin secretory granule biogenesis in metabolically stressed β cells
- PMID: 25157818
- PMCID: PMC4191024
- DOI: 10.1172/JCI74072
SORCS1 is necessary for normal insulin secretory granule biogenesis in metabolically stressed β cells
Abstract
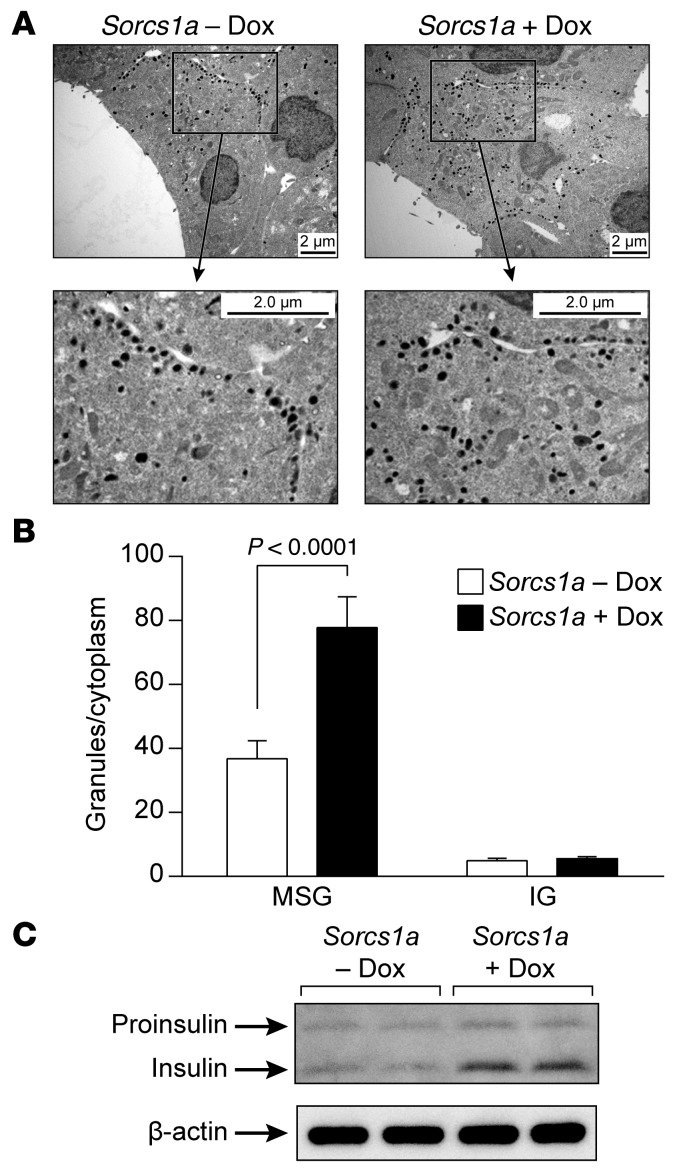
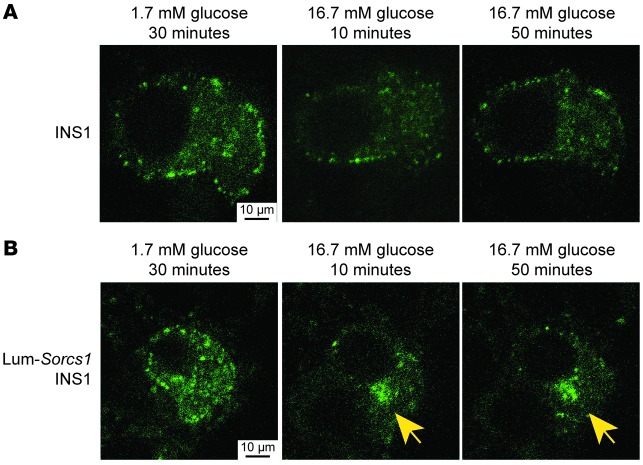
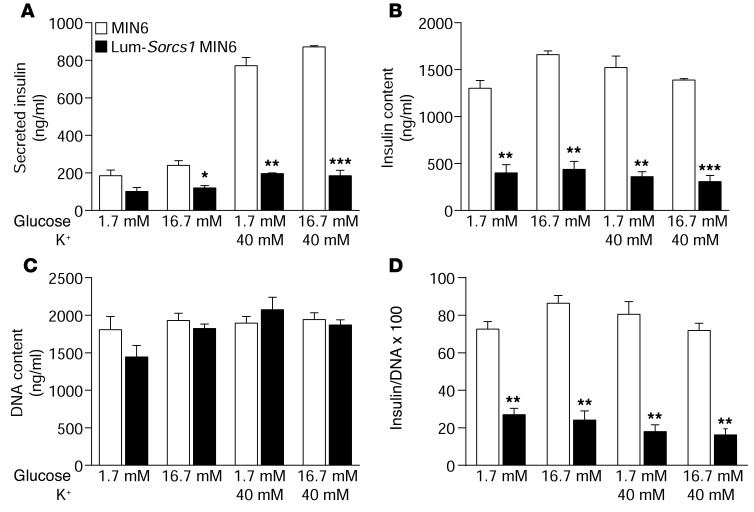
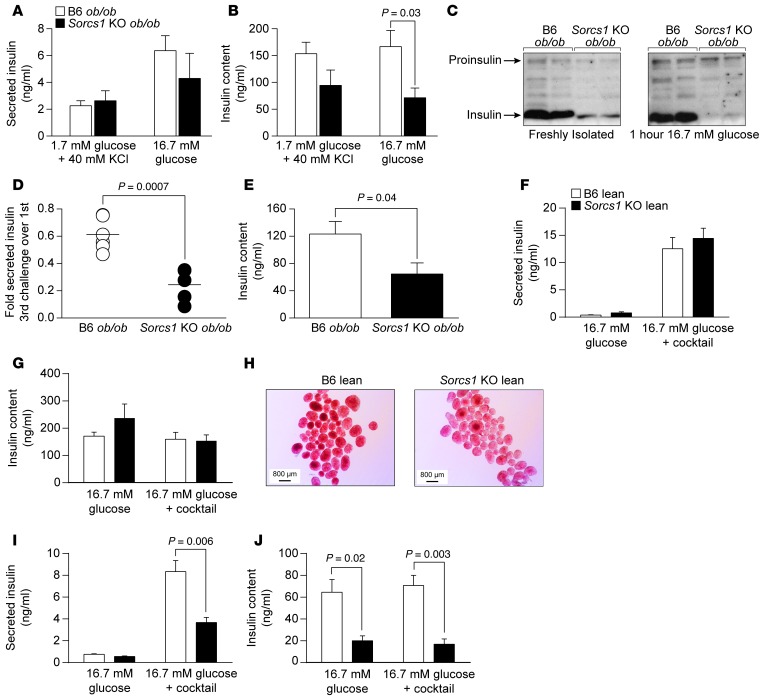
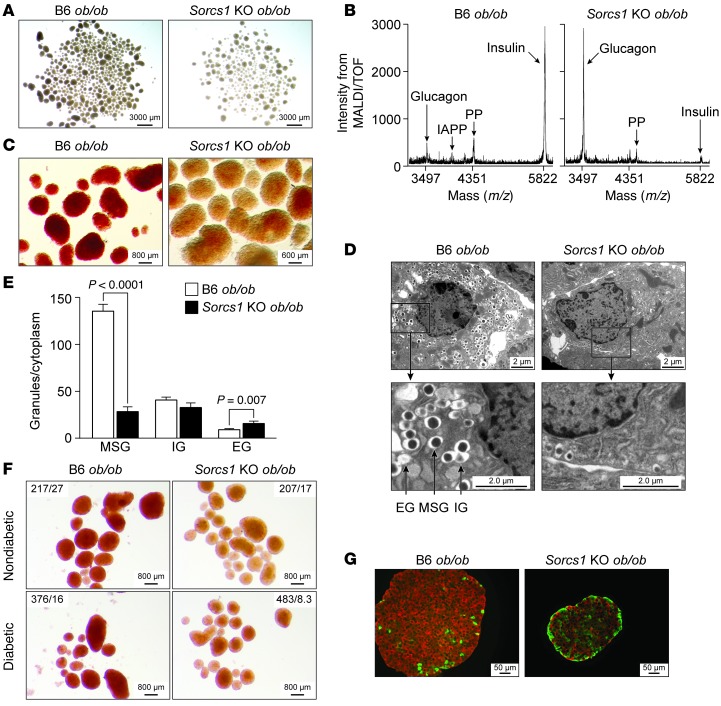
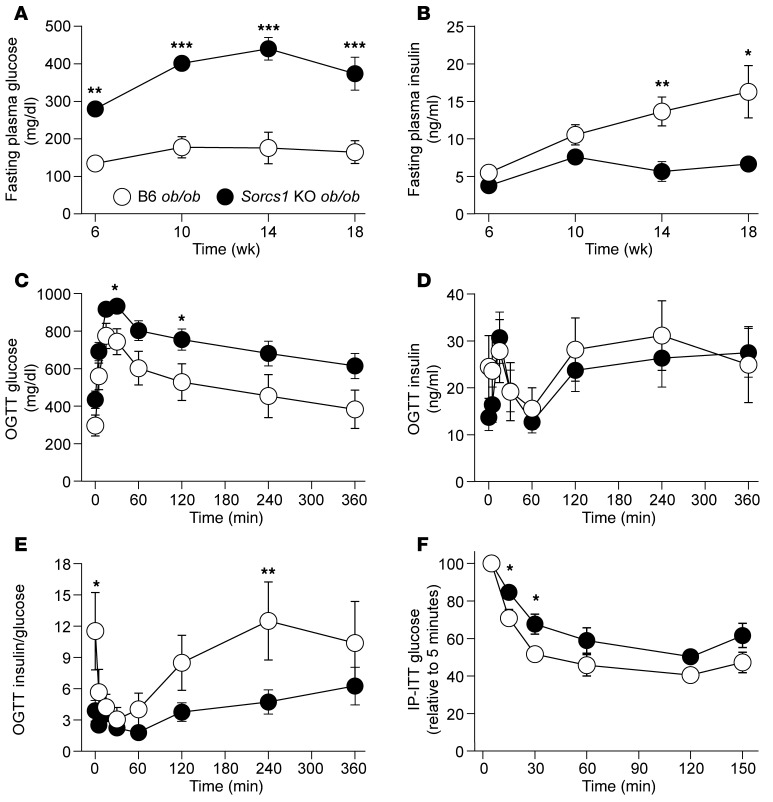
We previously positionally cloned Sorcs1 as a diabetes quantitative trait locus. Sorcs1 belongs to the Vacuolar protein sorting-10 (Vps10) gene family. In yeast, Vps10 transports enzymes from the trans-Golgi network (TGN) to the vacuole. Whole-body Sorcs1 KO mice, when made obese with the leptin(ob) mutation (ob/ob), developed diabetes. β Cells from these mice had a severe deficiency of secretory granules (SGs) and insulin. Interestingly, a single secretagogue challenge failed to consistently elicit an insulin secretory dysfunction. However, multiple challenges of the Sorcs1 KO ob/ob islets consistently revealed an insulin secretion defect. The luminal domain of SORCS1 (Lum-Sorcs1), when expressed in a β cell line, acted as a dominant-negative, leading to SG and insulin deficiency. Using syncollin-dsRed5TIMER adenovirus, we found that the loss of Sorcs1 function greatly impairs the rapid replenishment of SGs following secretagogue challenge. Chronic exposure of islets from lean Sorcs1 KO mice to high glucose and palmitate depleted insulin content and evoked an insulin secretion defect. Thus, in metabolically stressed mice, Sorcs1 is important for SG replenishment, and under chronic challenge by insulin secretagogues, loss of Sorcs1 leads to diabetes. Overexpression of full-length SORCS1 led to a 2-fold increase in SG content, suggesting that SORCS1 is sufficient to promote SG biogenesis.
Figures
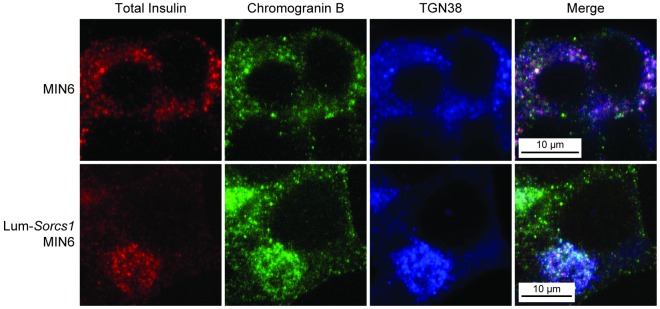
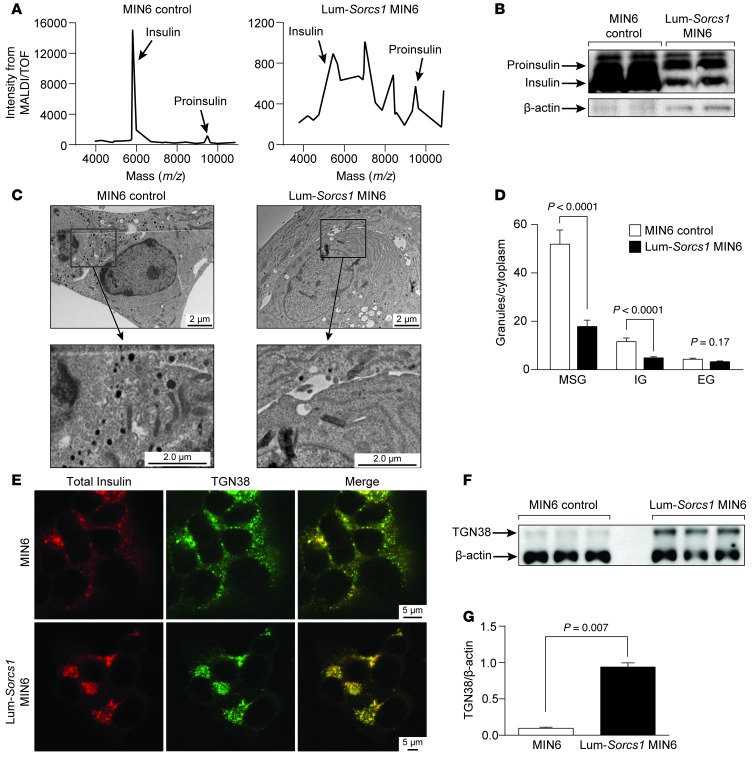
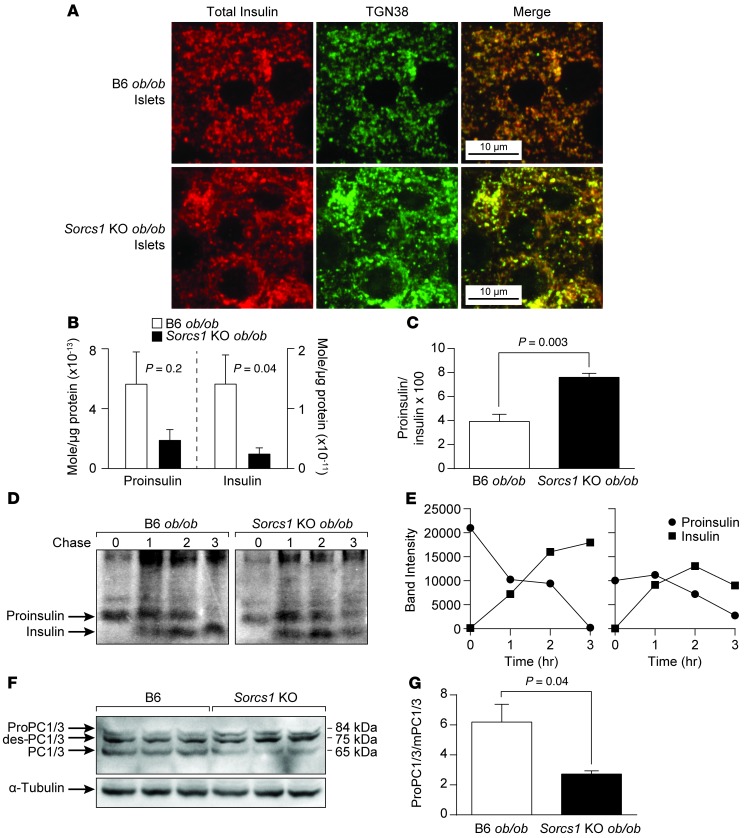
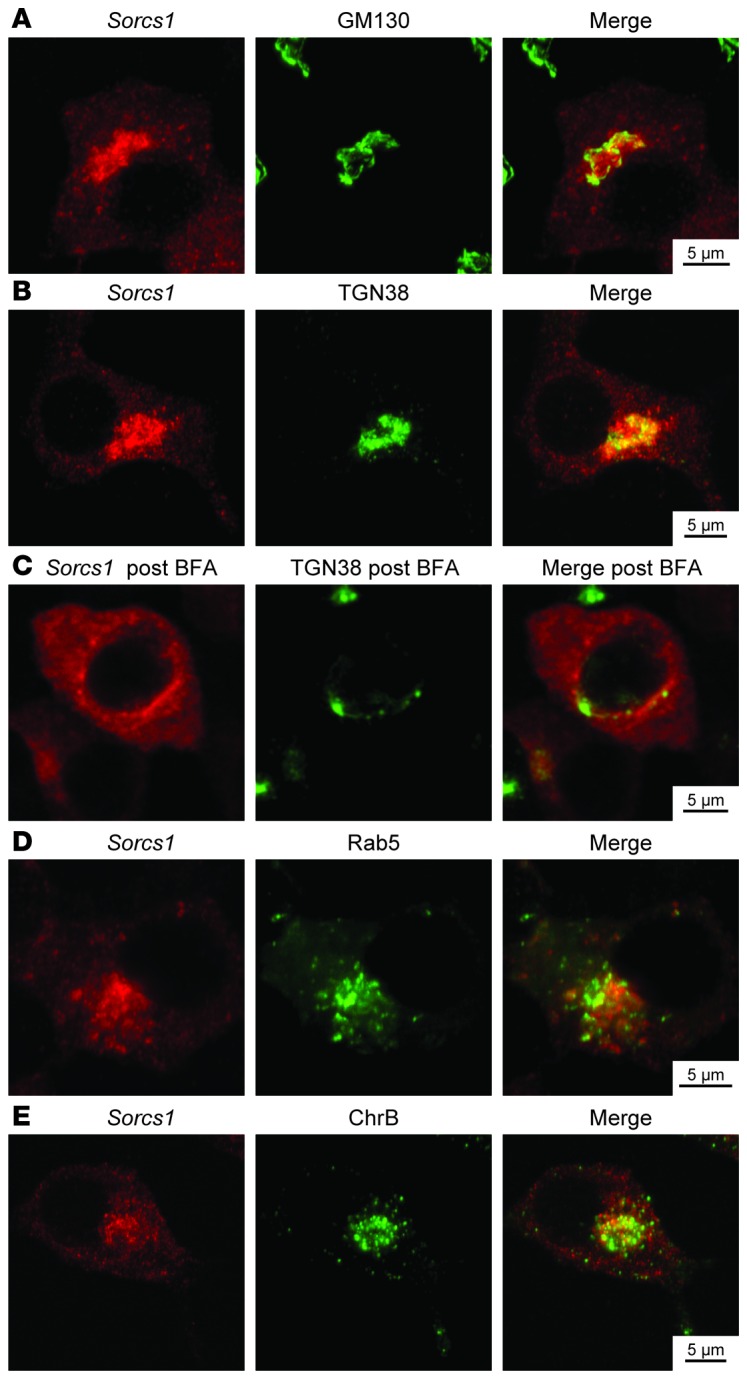
References
-
- Groop L, Lyssenko V. Genetic basis of β-cell dysfunction in man. Diabetes Obes Metab. 2009;11(suppl 4):149–158. - PubMed
-
- Benjannet S, Reudelhuber T, Mercure C, Rondeau N, Chretien M, Seidah NG. Proprotein conversion is determined by a multiplicity of factors including convertase processing, substrate specificity, and intracellular environment. Cell type-specific processing of human prorenin by the convertase PC1. J Biol Chem. 1992;267(16):11417–11423. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
