

A dopamine receptor contributes to paraquat-induced neurotoxicity in Drosophila
- PMID: 25158689
- PMCID: PMC4326327
- DOI: 10.1093/hmg/ddu430
A dopamine receptor contributes to paraquat-induced neurotoxicity in Drosophila
Abstract
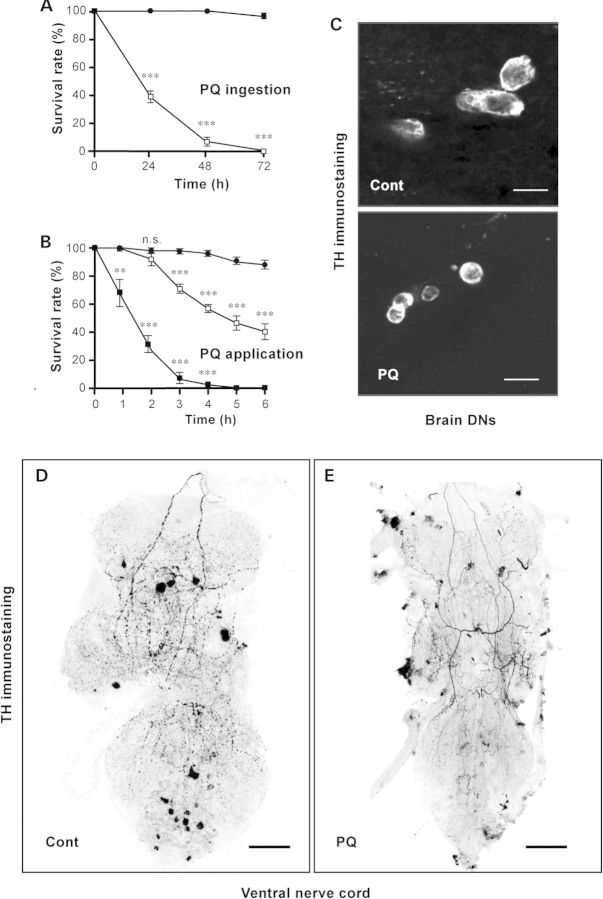
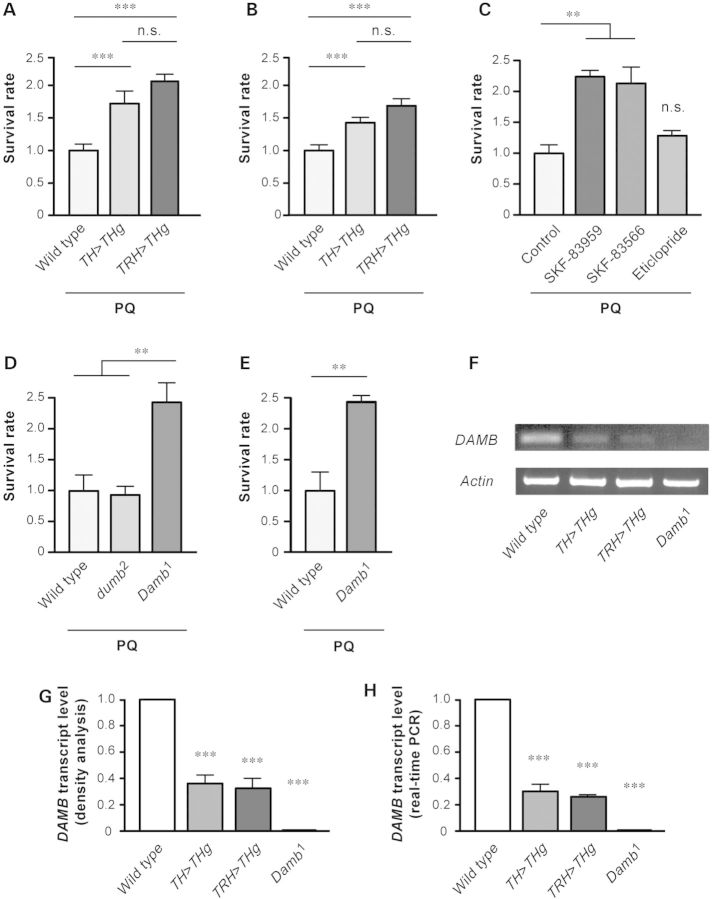
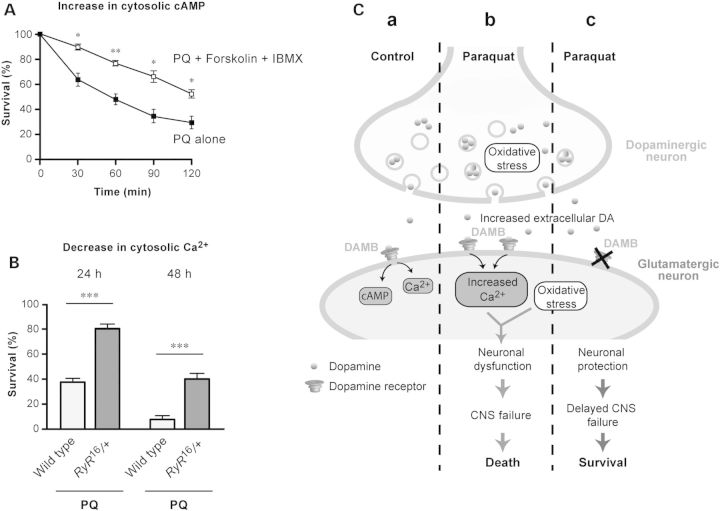
Long-term exposure to environmental oxidative stressors, like the herbicide paraquat (PQ), has been linked to the development of Parkinson's disease (PD), the most frequent neurodegenerative movement disorder. Paraquat is thus frequently used in the fruit fly Drosophila melanogaster and other animal models to study PD and the degeneration of dopaminergic neurons (DNs) that characterizes this disease. Here, we show that a D1-like dopamine (DA) receptor, DAMB, actively contributes to the fast central nervous system (CNS) failure induced by PQ in the fly. First, we found that a long-term increase in neuronal DA synthesis reduced DAMB expression and protected against PQ neurotoxicity. Secondly, a striking age-related decrease in PQ resistance in young adult flies correlated with an augmentation of DAMB expression. This aging-associated increase in oxidative stress vulnerability was not observed in a DAMB-deficient mutant. Thirdly, targeted inactivation of this receptor in glutamatergic neurons (GNs) markedly enhanced the survival of Drosophila exposed to either PQ or neurotoxic levels of DA, whereas, conversely, DAMB overexpression in these cells made the flies more vulnerable to both compounds. Fourthly, a mutation in the Drosophila ryanodine receptor (RyR), which inhibits activity-induced increase in cytosolic Ca(2+), also strongly enhanced PQ resistance. Finally, we found that DAMB overexpression in specific neuronal populations arrested development of the fly and that in vivo stimulation of either DNs or GNs increased PQ susceptibility. This suggests a model for DA receptor-mediated potentiation of PQ-induced neurotoxicity. Further studies of DAMB signaling in Drosophila could have implications for better understanding DA-related neurodegenerative disorders in humans.
© The Author 2014. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures




References
-
- Tsutsui H., Kinugawa S., Matsushima S. Oxidative stress and heart failure. Am. J. Physiol. Heart Circ. Physiol. 2011;301:H2181–H2190. - PubMed
-
- Lin M.T., Beal M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature. 2006;443:787–795. - PubMed
-
- Karbowski M., Neutzner A. Neurodegeneration as a consequence of failed mitochondrial maintenance. Acta Neuropathol. 2012;123:157–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
