

PLIN5 deletion remodels intracellular lipid composition and causes insulin resistance in muscle
- PMID: 25161888
- PMCID: PMC4142393
- DOI: 10.1016/j.molmet.2014.06.002
PLIN5 deletion remodels intracellular lipid composition and causes insulin resistance in muscle
Abstract
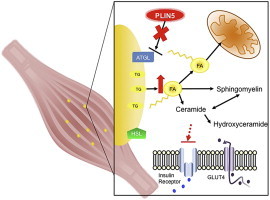
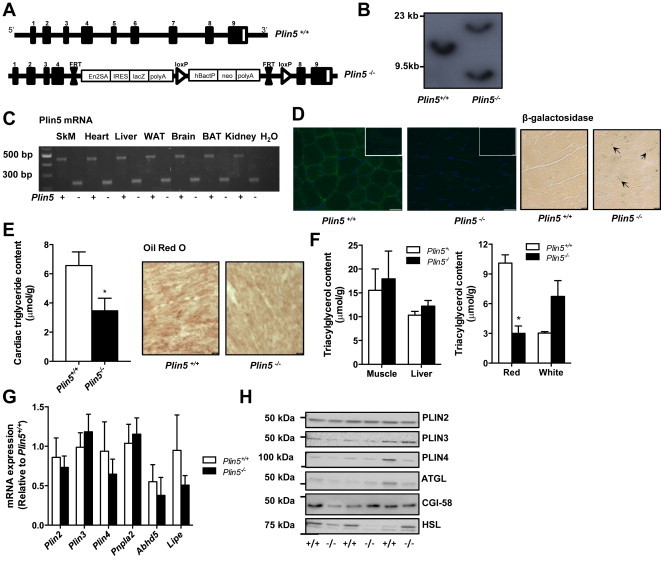
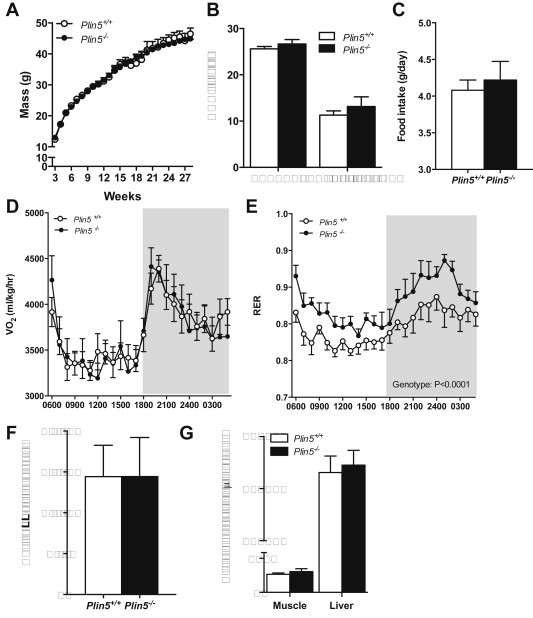
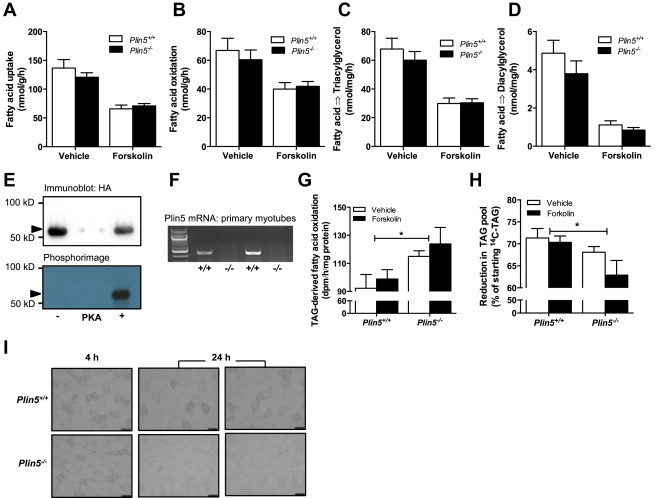
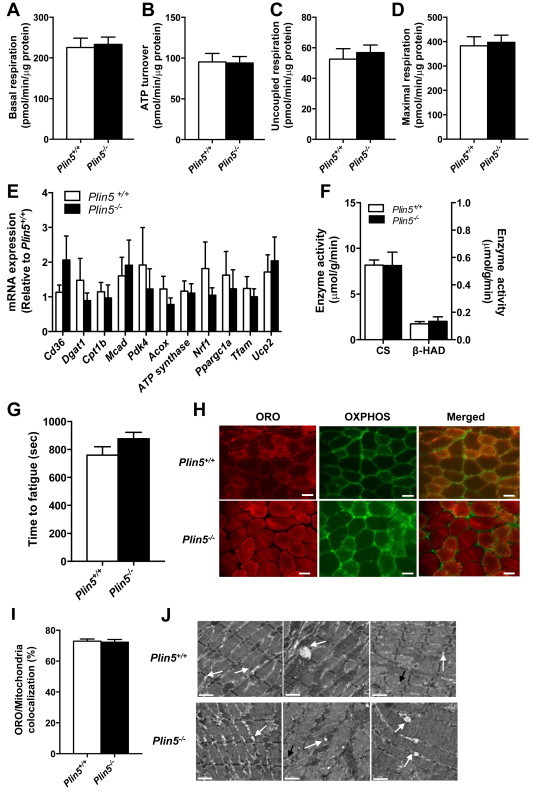
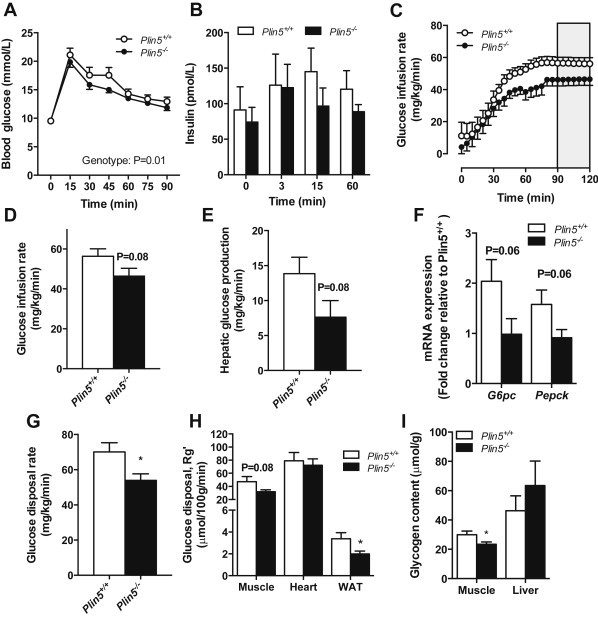
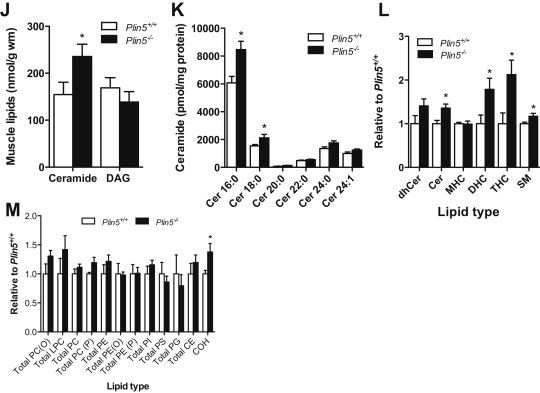
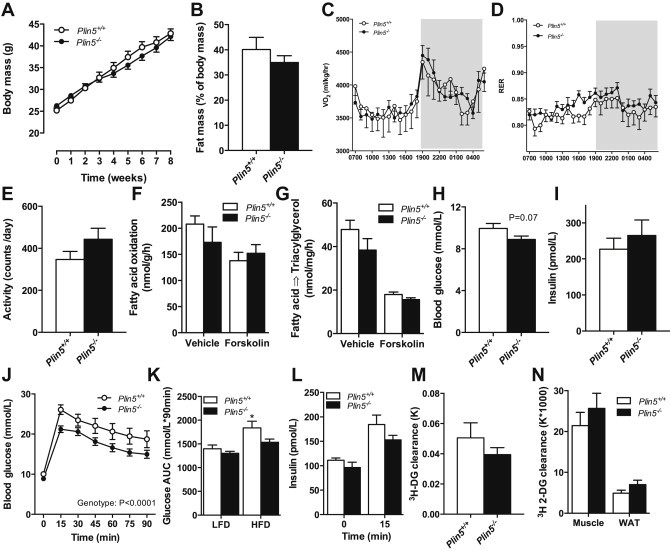
Defective control of lipid metabolism leading to lipotoxicity causes insulin resistance in skeletal muscle, a major factor leading to diabetes. Here, we demonstrate that perilipin (PLIN) 5 is required to couple intramyocellular triacylglycerol lipolysis with the metabolic demand for fatty acids. PLIN5 ablation depleted triacylglycerol stores but increased sphingolipids including ceramide, hydroxylceramides and sphingomyelin. We generated perilipin 5 (Plin5)(-/-) mice to determine the functional significance of PLIN5 in metabolic control and insulin action. Loss of PLIN5 had no effect on body weight, feeding or adiposity but increased whole-body carbohydrate oxidation. Plin5 (-/-) mice developed skeletal muscle insulin resistance, which was associated with ceramide accumulation. Liver insulin sensitivity was improved in Plin5 (-/-) mice, indicating tissue-specific effects of PLIN5 on insulin action. We conclude that PLIN5 plays a critical role in coordinating skeletal muscle triacylglycerol metabolism, which impacts sphingolipid metabolism, and is requisite for the maintenance of skeletal muscle insulin action.
Keywords: Insulin resistance; Lipid droplet; Lipid metabolism; Perilipin; Skeletal muscle.
Figures
Similar articles
-
Perilipin 5 fine-tunes lipid oxidation to metabolic demand and protects against lipotoxicity in skeletal muscle.Sci Rep. 2016 Dec 6;6:38310. doi: 10.1038/srep38310. Sci Rep. 2016. PMID: 27922115 Free PMC article.
-
Perilipin 5 Deletion in Hepatocytes Remodels Lipid Metabolism and Causes Hepatic Insulin Resistance in Mice.Diabetes. 2019 Mar;68(3):543-555. doi: 10.2337/db18-0670. Epub 2019 Jan 7. Diabetes. 2019. PMID: 30617219
-
Overexpression of PLIN5 in skeletal muscle promotes oxidative gene expression and intramyocellular lipid content without compromising insulin sensitivity.Biochim Biophys Acta. 2013 Apr;1831(4):844-52. doi: 10.1016/j.bbalip.2013.01.007. Epub 2013 Jan 22. Biochim Biophys Acta. 2013. PMID: 23353597
-
Perilipin 5, a lipid droplet protein adapted to mitochondrial energy utilization.Curr Opin Lipidol. 2014 Apr;25(2):110-7. doi: 10.1097/MOL.0000000000000057. Curr Opin Lipidol. 2014. PMID: 24535284 Free PMC article. Review.
-
Unraveling the roles of PLIN5: linking cell biology to physiology.Trends Endocrinol Metab. 2015 Mar;26(3):144-52. doi: 10.1016/j.tem.2015.01.005. Epub 2015 Feb 11. Trends Endocrinol Metab. 2015. PMID: 25682370 Review.
Cited by
-
Lipid droplet quantification based on iterative image processing.J Lipid Res. 2019 Jul;60(7):1333-1344. doi: 10.1194/jlr.D092841. Epub 2019 Mar 29. J Lipid Res. 2019. PMID: 30926625 Free PMC article.
-
Insulin resistance and the role of gamma-aminobutyric acid.J Res Med Sci. 2021 Jun 30;26:39. doi: 10.4103/jrms.JRMS_374_20. eCollection 2021. J Res Med Sci. 2021. PMID: 34484371 Free PMC article. Review.
-
Intramuscular triglyceride synthesis: importance in muscle lipid partitioning in humans.Am J Physiol Endocrinol Metab. 2018 Feb 1;314(2):E152-E164. doi: 10.1152/ajpendo.00142.2017. Epub 2017 Oct 3. Am J Physiol Endocrinol Metab. 2018. PMID: 28978544 Free PMC article.
-
Liver-derived extracellular vesicles improve whole-body glycaemic control via inter-organ communication.Nat Metab. 2024 Feb;6(2):254-272. doi: 10.1038/s42255-023-00971-z. Epub 2024 Jan 23. Nat Metab. 2024. PMID: 38263317
-
Perilipin5 protects against lipotoxicity and alleviates endoplasmic reticulum stress in pancreatic β-cells.Nutr Metab (Lond). 2019 Jul 30;16:50. doi: 10.1186/s12986-019-0375-2. eCollection 2019. Nutr Metab (Lond). 2019. PMID: 31384284 Free PMC article.
References
-
- Krssak M., Falk P.K., Dresner A., DiPietro L., Vogel S.M., Rothman D.L. Diabetologia. 1999;42:113–116. - PubMed
-
- Pan D.A., Lillioja S., Kriketos A.D., Milner M.R., Baur L.A., Bogardus C. Diabetes. 1997;46:983–988. - PubMed
-
- Perseghin G., Scifo P., De Cobelli F., Pagliato E., Battezzati A., Arcelloni C. Diabetes. 1999;48:1600–1606. - PubMed
-
- Jacob S., Machann J., Rett K., Brechtel K., Volk A., Renn W. Diabetes. 1999;48:1113–1119. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
