

Comparative analysis of metazoan chromatin organization
- PMID: 25164756
- PMCID: PMC4227084
- DOI: 10.1038/nature13415
Comparative analysis of metazoan chromatin organization
Abstract
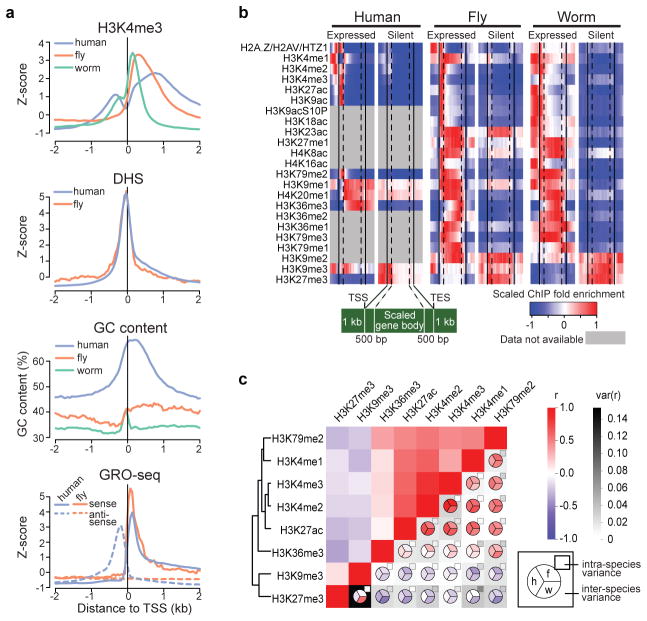
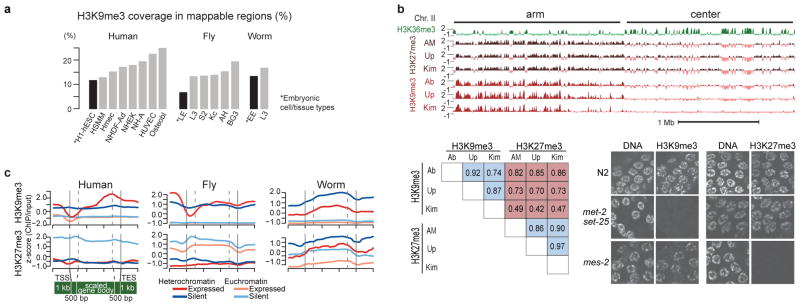
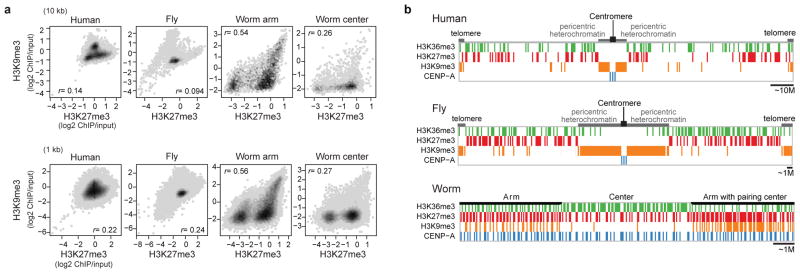
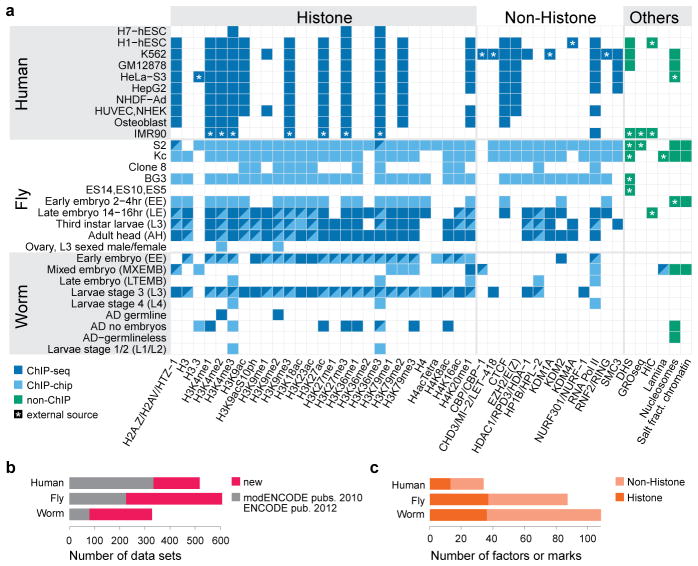
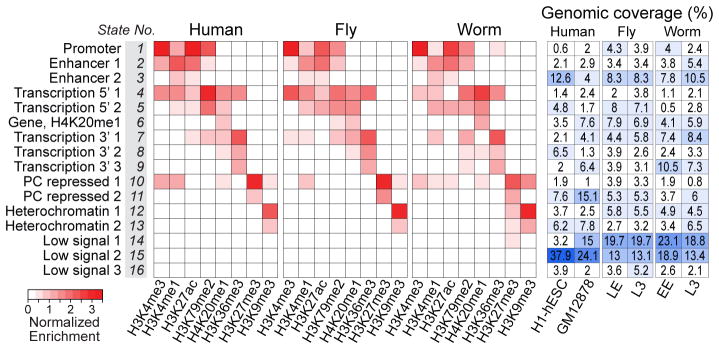
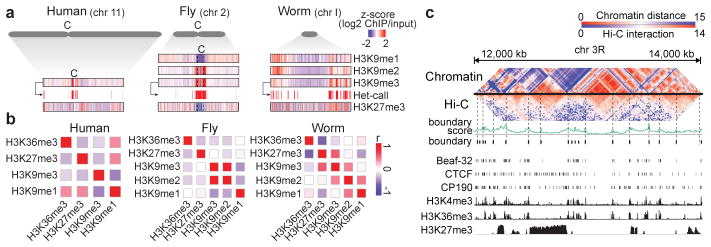
Genome function is dynamically regulated in part by chromatin, which consists of the histones, non-histone proteins and RNA molecules that package DNA. Studies in Caenorhabditis elegans and Drosophila melanogaster have contributed substantially to our understanding of molecular mechanisms of genome function in humans, and have revealed conservation of chromatin components and mechanisms. Nevertheless, the three organisms have markedly different genome sizes, chromosome architecture and gene organization. On human and fly chromosomes, for example, pericentric heterochromatin flanks single centromeres, whereas worm chromosomes have dispersed heterochromatin-like regions enriched in the distal chromosomal 'arms', and centromeres distributed along their lengths. To systematically investigate chromatin organization and associated gene regulation across species, we generated and analysed a large collection of genome-wide chromatin data sets from cell lines and developmental stages in worm, fly and human. Here we present over 800 new data sets from our ENCODE and modENCODE consortia, bringing the total to over 1,400. Comparison of combinatorial patterns of histone modifications, nuclear lamina-associated domains, organization of large-scale topological domains, chromatin environment at promoters and enhancers, nucleosome positioning, and DNA replication patterns reveals many conserved features of chromatin organization among the three organisms. We also find notable differences in the composition and locations of repressive chromatin. These data sets and analyses provide a rich resource for comparative and species-specific investigations of chromatin composition, organization and function.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Genomics: Hiding in plain sight.Nature. 2014 Aug 28;512(7515):374-5. doi: 10.1038/512374a. Nature. 2014. PMID: 25164742 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- 5RL9EB008539/EB/NIBIB NIH HHS/United States
- U54 HG006991/HG/NHGRI NIH HHS/United States
- R01 HG004037/HG/NHGRI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- U01 HG004279/HG/NHGRI NIH HHS/United States
- R37 GM048405/GM/NIGMS NIH HHS/United States
- U54 CA121852/CA/NCI NIH HHS/United States
- U54 HG004570/HG/NHGRI NIH HHS/United States
- T32 GM071340/GM/NIGMS NIH HHS/United States
- 101863/WT_/Wellcome Trust/United Kingdom
- U54HG004570/HG/NHGRI NIH HHS/United States
- K99HG006259/HG/NHGRI NIH HHS/United States
- R01 GM098461/GM/NIGMS NIH HHS/United States
- 54523/WT_/Wellcome Trust/United Kingdom
- U54CA121852/CA/NCI NIH HHS/United States
- U01HG004270/HG/NHGRI NIH HHS/United States
- 092096/WT_/Wellcome Trust/United Kingdom
- T32 HG002295/HG/NHGRI NIH HHS/United States
- U01HG004279/HG/NHGRI NIH HHS/United States
- U01HG004695/HG/NHGRI NIH HHS/United States
- RL9 EB008539/EB/NIBIB NIH HHS/United States
- U01HG004258/HG/NHGRI NIH HHS/United States
- U01 HG004258/HG/NHGRI NIH HHS/United States
- U01 HG004270/HG/NHGRI NIH HHS/United States
- U01 HG004695/HG/NHGRI NIH HHS/United States
- K99 HG006259/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
