

Distinct mechanisms regulate exposure of neutralizing epitopes in the V2 and V3 loops of HIV-1 envelope
- PMID: 25165106
- PMCID: PMC4248937
- DOI: 10.1128/JVI.02125-14
Distinct mechanisms regulate exposure of neutralizing epitopes in the V2 and V3 loops of HIV-1 envelope
Abstract
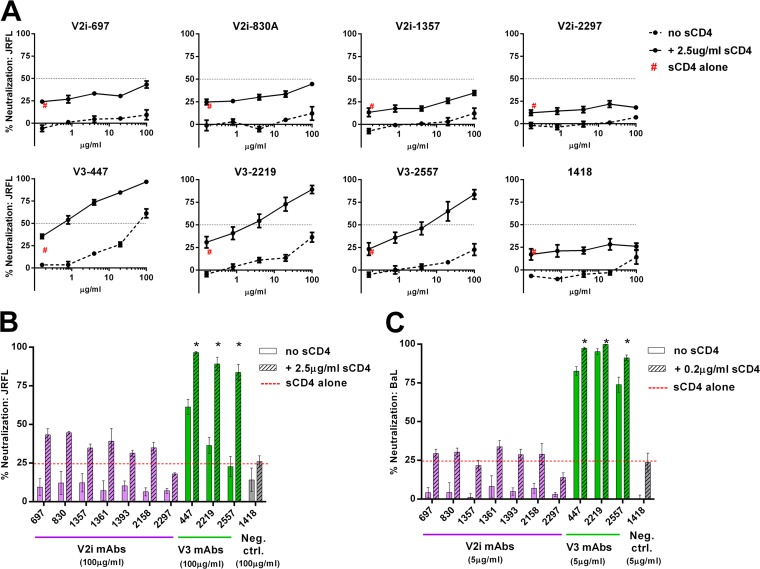
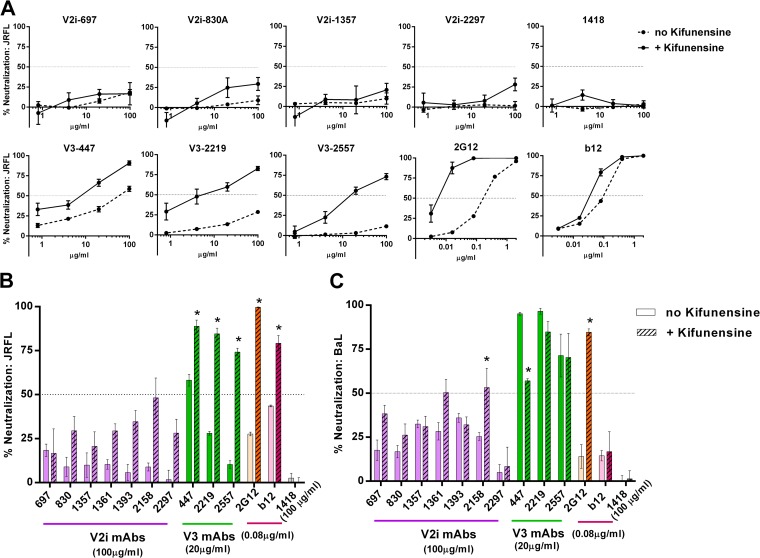
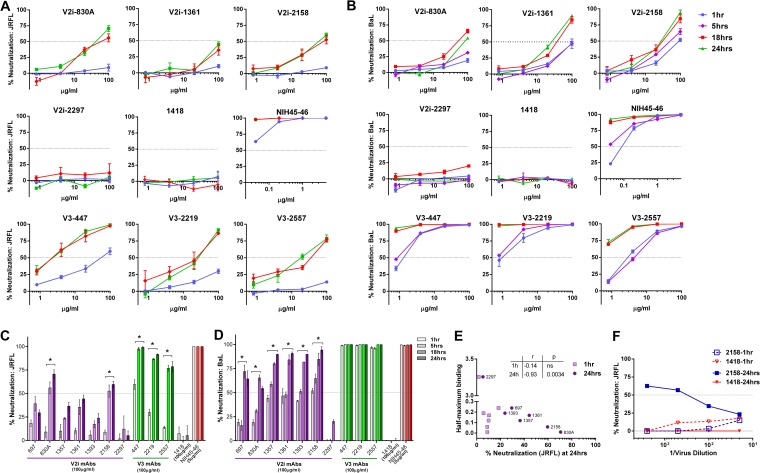
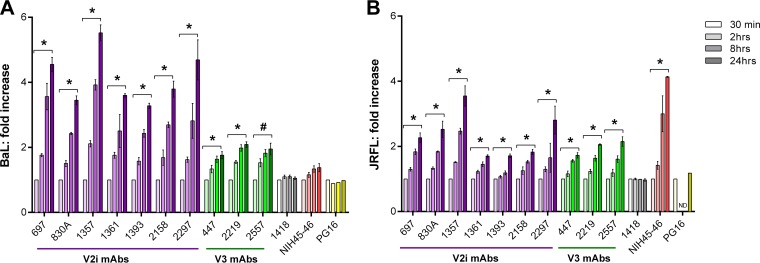
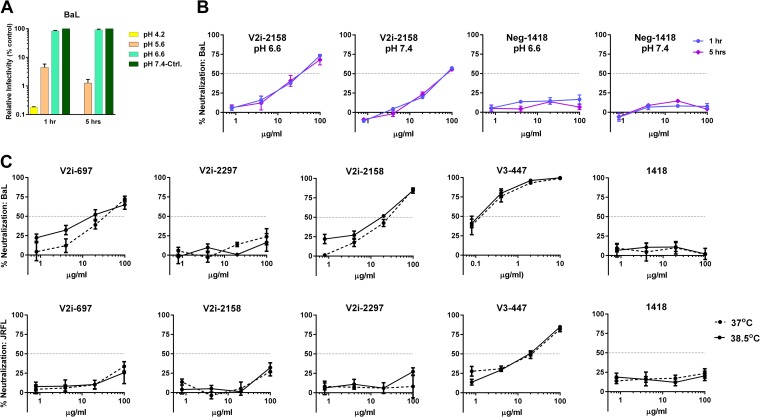
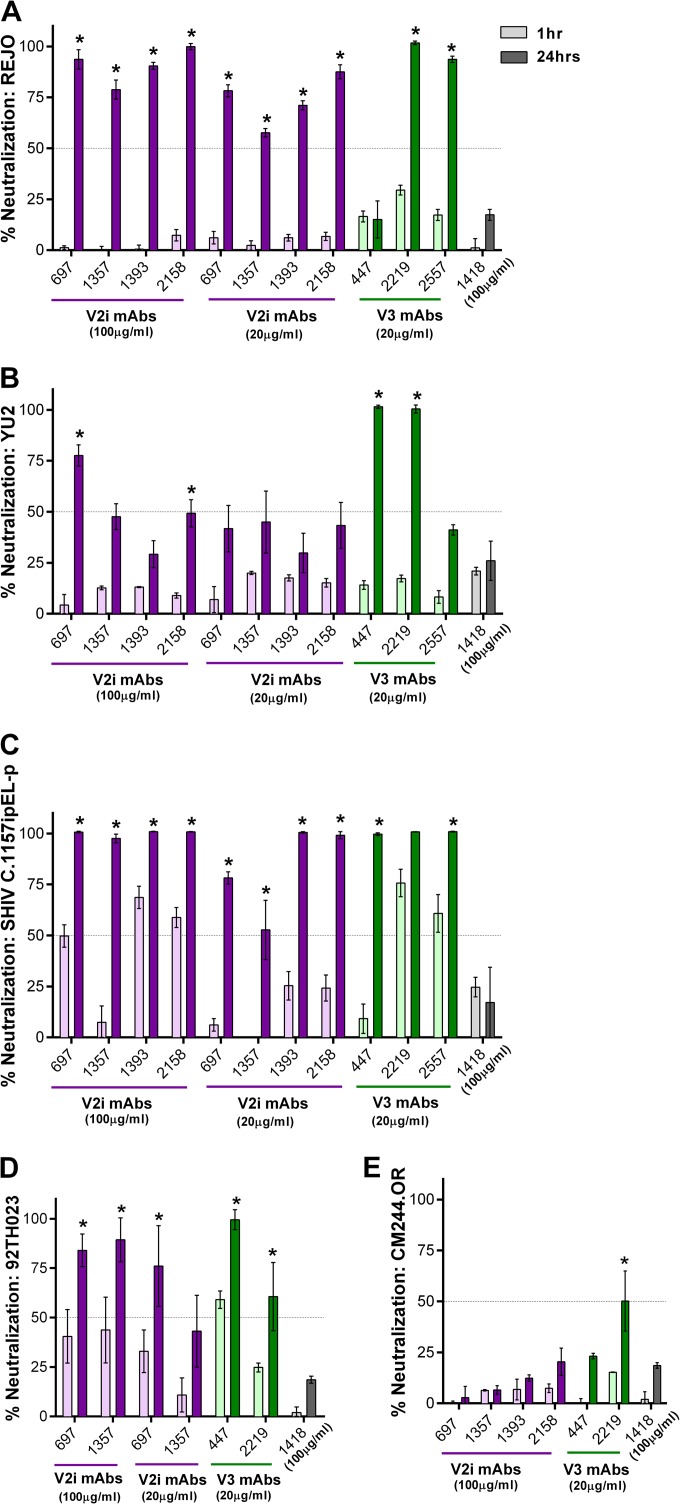
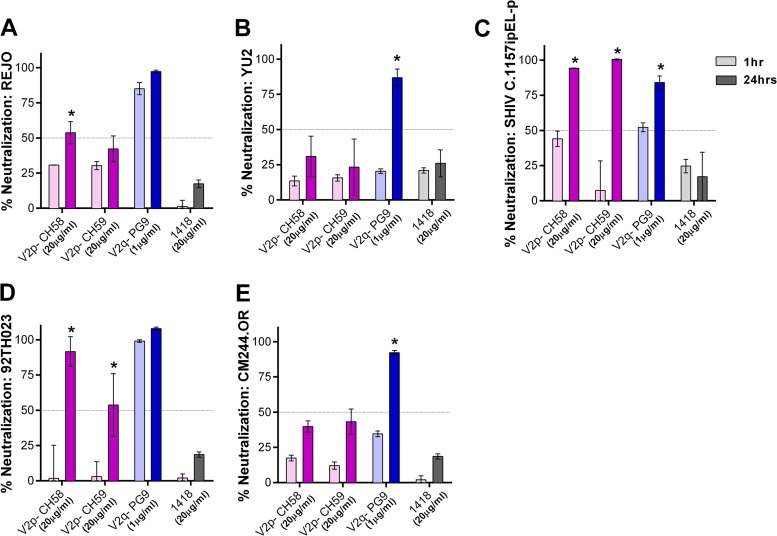
Broadly neutralizing antibodies targeting the HIV-1 envelope (Env) are key components for protection against HIV-1. However, many cross-reactive epitopes are often occluded. This study investigates the mechanisms contributing to the masking of V2i (variable loop V2 integrin) epitopes compared to the accessibility of V3 epitopes. V2i are conformation-dependent epitopes encompassing the integrin α4β7-binding motif on the V1V2 loop of HIV-1 Env gp120. The V2i monoclonal antibodies (MAbs) display extensive cross-reactivity with gp120 monomers from many subtypes but neutralize only few viruses, indicating V2i's cryptic nature. First, we asked whether CD4-induced Env conformational changes affect V2i epitopes similarly to V3. CD4 treatment of BaL and JRFL pseudoviruses increased their neutralization sensitivity to V3 MAbs but not to the V2i MAbs. Second, the contribution of N-glycans in masking V2i versus V3 epitopes was evaluated by testing the neutralization of pseudoviruses produced in the presence of a glycosidase inhibitor, kifunensine. Viruses grown in kifunensine were more sensitive to neutralization by V3 but not V2i MAbs. Finally, we evaluated the time-dependent dynamics of the V2i and V3 epitopes. Extending the time of virus-MAb interaction to 18 h before adding target cells increased virus neutralization by some V2i MAbs and all V3 MAbs tested. Consistent with this, V2i MAb binding to Env on the surface of transfected cells also increased in a time-dependent manner. Hence, V2i and V3 epitopes are highly dynamic, but distinct factors modulate the antibody accessibility of these epitopes. The study reveals the importance of the structural dynamics of V2i and V3 epitopes in determining HIV-1 neutralization by antibodies targeting these sites.
Importance: Conserved neutralizing epitopes are present in the V1V2 and V3 regions of HIV-1 Env, but these epitopes are often occluded from Abs. This study reveals that distinct mechanisms contribute to the masking of V3 epitopes and V2i epitopes in the V1V2 domain. Importantly, V3 MAbs and some V2i MAbs display greater neutralization against relatively resistant HIV-1 isolates when the MAbs interact with the virus for a prolonged period of time. Given their highly immunogenic nature, V3 and V2i epitopes are valuable targets that would augment the efficacy of HIV vaccines.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Modulation of Antibody Responses to the V1V2 and V3 Regions of HIV-1 Envelope by Immune Complex Vaccines.Front Immunol. 2018 Oct 26;9:2441. doi: 10.3389/fimmu.2018.02441. eCollection 2018. Front Immunol. 2018. PMID: 30416503 Free PMC article.
-
Rationally Targeted Mutations at the V1V2 Domain of the HIV-1 Envelope to Augment Virus Neutralization by Anti-V1V2 Monoclonal Antibodies.PLoS One. 2015 Oct 22;10(10):e0141233. doi: 10.1371/journal.pone.0141233. eCollection 2015. PLoS One. 2015. PMID: 26491873 Free PMC article.
-
Plasticity and Epitope Exposure of the HIV-1 Envelope Trimer.J Virol. 2017 Aug 10;91(17):e00410-17. doi: 10.1128/JVI.00410-17. Print 2017 Sep 1. J Virol. 2017. PMID: 28615206 Free PMC article.
-
Vaccine-induced V1V2-specific antibodies control and or protect against infection with HIV, SIV and SHIV.Curr Opin HIV AIDS. 2019 Jul;14(4):309-317. doi: 10.1097/COH.0000000000000551. Curr Opin HIV AIDS. 2019. PMID: 30994501 Free PMC article. Review.
-
Specificity of the autologous neutralizing antibody response.Curr Opin HIV AIDS. 2009 Sep;4(5):358-63. doi: 10.1097/COH.0b013e32832ea7e8. Curr Opin HIV AIDS. 2009. PMID: 20048698 Free PMC article. Review.
Cited by
-
HIV-1 Envelope Glycan Composition as a Key Determinant of Efficient Virus Transmission via DC-SIGN and Resistance to Inhibitory Lectins.iScience. 2019 Nov 22;21:413-427. doi: 10.1016/j.isci.2019.10.030. Epub 2019 Oct 18. iScience. 2019. PMID: 31704652 Free PMC article.
-
HIV vaccine delayed boosting increases Env variable region 2-specific antibody effector functions.JCI Insight. 2020 Jan 30;5(2):e131437. doi: 10.1172/jci.insight.131437. JCI Insight. 2020. PMID: 31996483 Free PMC article.
-
Alterations of HIV-1 envelope phenotype and antibody-mediated neutralization by signal peptide mutations.PLoS Pathog. 2018 Jan 25;14(1):e1006812. doi: 10.1371/journal.ppat.1006812. eCollection 2018 Jan. PLoS Pathog. 2018. PMID: 29370305 Free PMC article.
-
Identification of New Regions in HIV-1 gp120 Variable 2 and 3 Loops that Bind to α4β7 Integrin Receptor.PLoS One. 2015 Dec 1;10(12):e0143895. doi: 10.1371/journal.pone.0143895. eCollection 2015. PLoS One. 2015. PMID: 26625359 Free PMC article. Clinical Trial.
-
Modulation of Antibody Responses to the V1V2 and V3 Regions of HIV-1 Envelope by Immune Complex Vaccines.Front Immunol. 2018 Oct 26;9:2441. doi: 10.3389/fimmu.2018.02441. eCollection 2018. Front Immunol. 2018. PMID: 30416503 Free PMC article.
References
-
- Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, Benenson M, Gurunathan S, Tartaglia J, McNeil JG, Francis DP, Stablein D, Birx DL, Chunsuttiwat S, Khamboonruang C, Thongcharoen P, Robb ML, Michael NL, Kunasol P, Kim JH. 2009. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 361:2209–2220. 10.1056/NEJMoa0908492. - DOI - PubMed
-
- Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH. 2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 366:1275–1286. 10.1056/NEJMoa1113425. - DOI - PMC - PubMed
-
- Zolla-Pazner S, Decamp A, Gilbert PB, Williams C, Yates NL, Williams WT, Howington R, Fong Y, Morris DE, Soderberg KA, Irene C, Reichman C, Pinter A, Parks R, Pitisuttithum P, Kaewkungwal J, Rerks-Ngarm S, Nitayaphan S, Andrews C, O'Connell RJ, Yang ZY, Nabel GJ, Kim JH, Michael NL, Montefiori DC, Liao HX, Haynes BF, Tomaras GD. 2014. Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLoS One 9:e87572. 10.1371/journal.pone.0087572. - DOI - PMC - PubMed
-
- Zolla-Pazner S, deCamp AC, Cardozo T, Karasavvas N, Gottardo R, Williams C, Morris DE, Tomaras G, Rao M, Billings E, Berman P, Shen X, Andrews C, O'Connell RJ, Ngauy V, Nitayaphan S, de Souza M, Korber B, Koup R, Bailer RT, Mascola JR, Pinter A, Montefiori D, Haynes BF, Robb ML, Rerks-Ngarm S, Michael NL, Gilbert PB, Kim JH. 2013. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PLoS One 8:e53629. 10.1371/journal.pone.0053629. - DOI - PMC - PubMed
-
- Rolland M, Edlefsen PT, Larsen BB, Tovanabutra S, Sanders-Buell E, Hertz T, deCamp AC, Carrico C, Menis S, Magaret CA, Ahmed H, Juraska M, Chen L, Konopa P, Nariya S, Stoddard JN, Wong K, Zhao H, Deng W, Maust BS, Bose M, Howell S, Bates A, Lazzaro M, O'Sullivan A, Lei E, Bradfield A, Ibitamuno G, Assawadarachai V, O'Connell RJ, deSouza MS, Nitayaphan S, Rerks-Ngarm S, Robb ML, McLellan JS, Georgiev I, Kwong PD, Carlson JM, Michael NL, Schief WR, Gilbert PB, Mullins JI, Kim JH. 2012. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature 490:417–420. 10.1038/nature11519. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
