

Structural definition of an antibody-dependent cellular cytotoxicity response implicated in reduced risk for HIV-1 infection
- PMID: 25165110
- PMCID: PMC4248932
- DOI: 10.1128/JVI.02194-14
Structural definition of an antibody-dependent cellular cytotoxicity response implicated in reduced risk for HIV-1 infection
Abstract
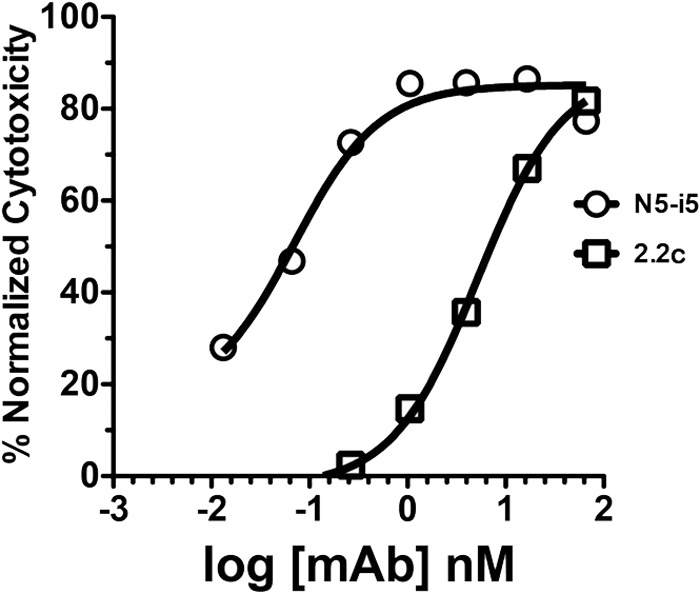
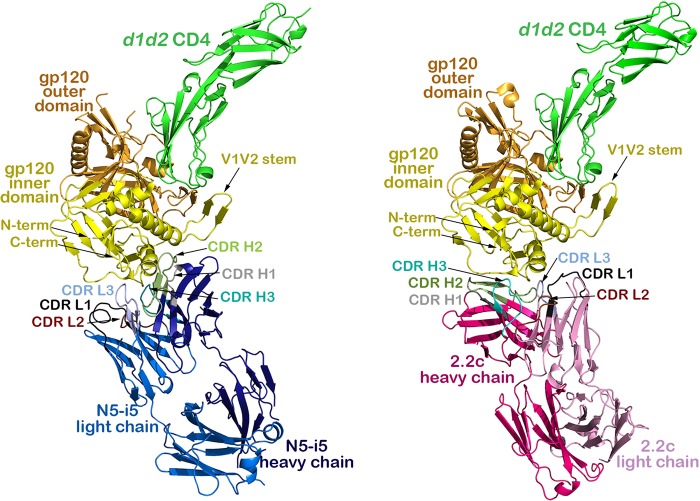
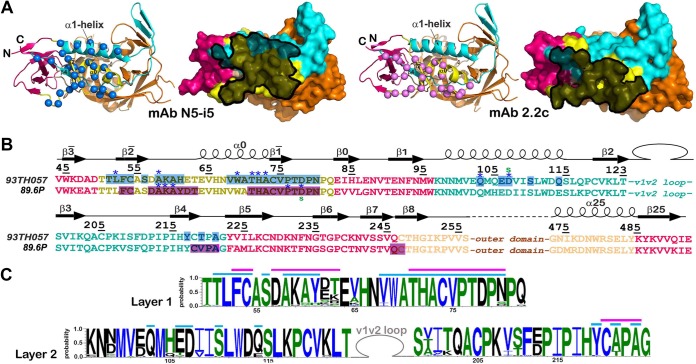
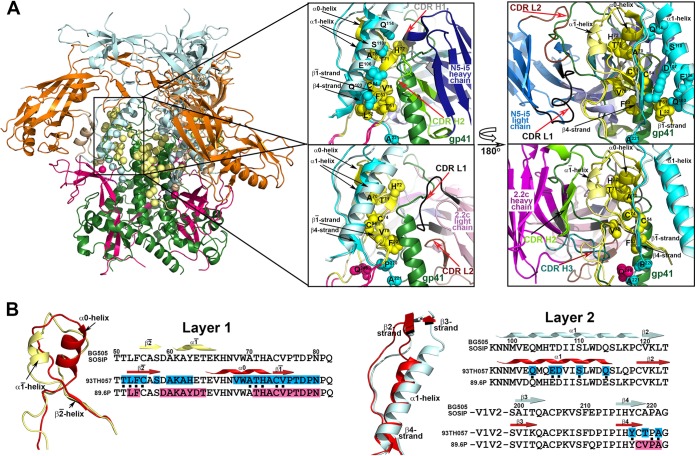
The RV144 vaccine trial implicated epitopes in the C1 region of gp120 (A32-like epitopes) as targets of potentially protective antibody-dependent cellular cytotoxicity (ADCC) responses. A32-like epitopes are highly immunogenic, as infected or vaccinated individuals frequently produce antibodies specific for these determinants. Antibody titers, as measured by enzyme-linked immunosorbent assay (ELISA) against these epitopes, however, do not consistently correlate with protection. Here, we report crystal structures of CD4-stabilized gp120 cores complexed with the Fab fragments of two nonneutralizing, A32-like monoclonal antibodies (MAbs), N5-i5 and 2.2c, that compete for antigen binding and have similar antigen-binding affinities yet exhibit a 75-fold difference in ADCC potency. We find that these MAbs recognize overlapping epitopes formed by mobile layers 1 and 2 of the gp120 inner domain, including the C1 and C2 regions, but bind gp120 at different angles via juxtaposed VH and VL contact surfaces. A comparison of structural and immunological data further showed that antibody orientation on bound antigen and the capacity to form multivalent antigen-antibody complexes on target cells were key determinants of ADCC potency, with the latter process having the greater impact. These studies provide atomic-level definition of A32-like epitopes implicated as targets of protective antibodies in RV144. Moreover, these studies establish that epitope structure and mode of antibody binding can dramatically affect the potency of Fc-mediated effector function against HIV-1. These results provide key insights for understanding, refining, and improving the outcome of HIV vaccine trials, in which relevant immune responses are facilitated by A32-like elicited responses.
Importance: HIV-1 Env is a primary target for antibodies elicited during infection. Although a small number of infected individuals elicit broadly neutralizing antibodies, the bulk of the humoral response consists of antibodies that do not neutralize or do so with limited breadth but may effect protection through Fc receptor-dependent processes, such as antibody-dependent cellular cytotoxicity (ADCC). Understanding these nonneutralizing responses is an important aspect of elucidating the complete spectrum of immune response against HIV-1 infection. With this report, we provide the first atomic-level definition of nonneutralizing CD4-induced epitopes in the N-terminal region of the HIV-1 gp120 (A32-like epitopes). Further, our studies point to the dominant role of precise epitope targeting and mode of antibody attachment in ADCC responses even when largely overlapping epitopes are involved. Such information provides key insights into the mechanisms of Fc-mediated function of antibodies to HIV-1 and will help us understand the outcome of vaccine trials based on humoral immunity.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
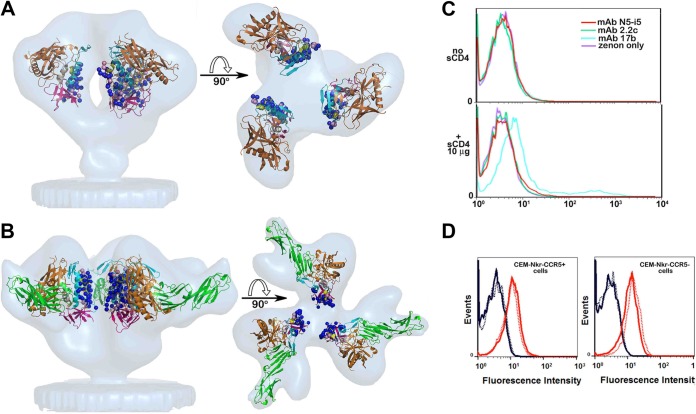
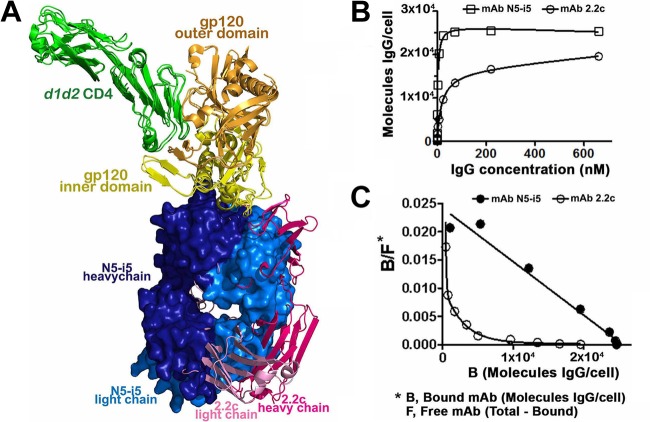
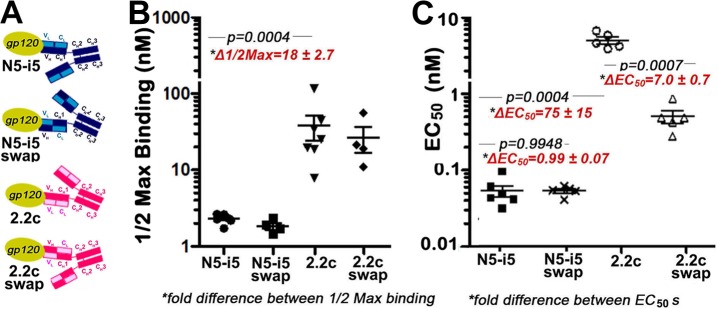
References
-
- Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH. 2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 366:1275–1286. 10.1056/NEJMoa1113425. - DOI - PMC - PubMed
-
- Alpert MD, Harvey JD, Lauer WA, Reeves RK, Piatak M, Jr, Carville A, Mansfield KG, Lifson JD, Li W, Desrosiers RC, Johnson RP, Evans DT. 2012. ADCC develops over time during persistent infection with live-attenuated SIV and is associated with complete protection against SIV(mac)251 challenge. PLoS Pathog. 8:e1002890. 10.1371/journal.ppat.1002890. - DOI - PMC - PubMed
-
- Lambotte O, Ferrari G, Moog C, Yates NL, Liao HX, Parks RJ, Hicks CB, Owzar K, Tomaras GD, Montefiori DC, Haynes BF, Delfraissy JF. 2009. Heterogeneous neutralizing antibody and antibody-dependent cell cytotoxicity responses in HIV-1 elite controllers. AIDS 23:897–906. 10.1097/QAD.0b013e328329f97d. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
