

Cell type-dependent RNA recombination frequency in the Japanese encephalitis virus
- PMID: 25165704
- PMCID: PMC4140105
- DOI: 10.1155/2014/471323
Cell type-dependent RNA recombination frequency in the Japanese encephalitis virus
Abstract
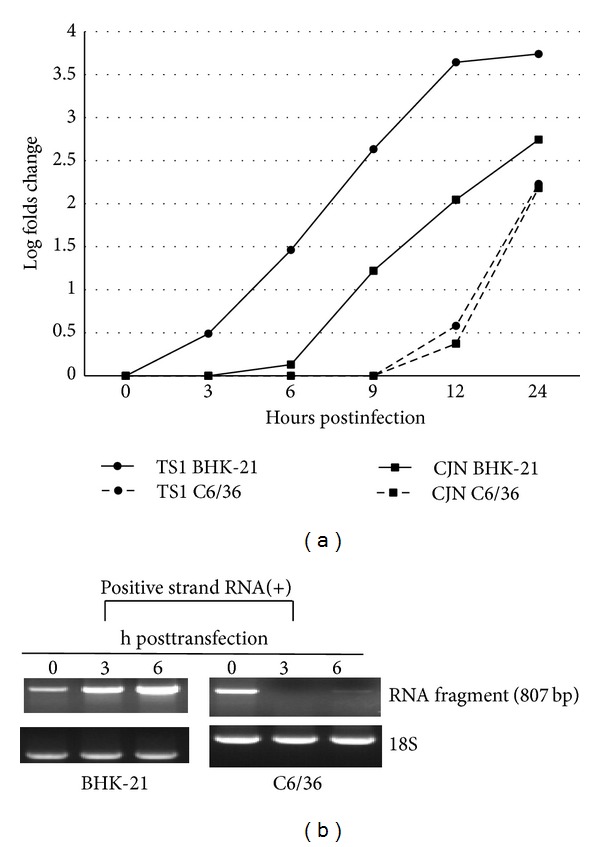
Japanese encephalitis virus (JEV) is one of approximately 70 flaviviruses, frequently causing symptoms involving the central nervous system. Mutations of its genomic RNA frequently occur during viral replication, which is believed to be a force contributing to viral evolution. Nevertheless, accumulating evidences show that some JEV strains may have actually arisen from RNA recombination between genetically different populations of the virus. We have demonstrated that RNA recombination in JEV occurs unequally in different cell types. In the present study, viral RNA fragments transfected into as well as viral RNAs synthesized in mosquito cells were shown not to be stable, especially in the early phase of infection possibly via cleavage by exoribonuclease. Such cleaved small RNA fragments may be further degraded through an RNA interference pathway triggered by viral double-stranded RNA during replication in mosquito cells, resulting in a lower frequency of RNA recombination in mosquito cells compared to that which occurs in mammalian cells. In fact, adjustment of viral RNA to an appropriately lower level in mosquito cells prevents overgrowth of the virus and is beneficial for cells to survive the infection. Our findings may also account for the slower evolution of arboviruses as reported previously.
Figures
Similar articles
-
Cellular DDX3 regulates Japanese encephalitis virus replication by interacting with viral un-translated regions.Virology. 2014 Jan 20;449:70-81. doi: 10.1016/j.virol.2013.11.008. Epub 2013 Nov 26. Virology. 2014. PMID: 24418539 Free PMC article.
-
Susceptibility of naïve and differentiated PC12 cells to Japanese encephalitis virus infection.IUBMB Life. 2017 Feb;69(2):79-87. doi: 10.1002/iub.1595. Epub 2017 Jan 23. IUBMB Life. 2017. PMID: 28111888
-
Viral Infection of the Central Nervous System and Neuroinflammation Precede Blood-Brain Barrier Disruption during Japanese Encephalitis Virus Infection.J Virol. 2015 May;89(10):5602-14. doi: 10.1128/JVI.00143-15. Epub 2015 Mar 11. J Virol. 2015. PMID: 25762733 Free PMC article.
-
Japanese encephalitis virus: from genome to infectome.Microbes Infect. 2011 Apr;13(4):312-21. doi: 10.1016/j.micinf.2011.01.002. Epub 2011 Jan 14. Microbes Infect. 2011. PMID: 21238600 Review.
-
Japanese encephalitis virus invasion of cell: allies and alleys.Rev Med Virol. 2016 Mar;26(2):129-41. doi: 10.1002/rmv.1868. Epub 2015 Dec 23. Rev Med Virol. 2016. PMID: 26695690 Review.
Cited by
-
Genetic diversity of Murray Valley encephalitis virus 1951-2020 identified via phylogenetic and evolutionary analyses.PLoS Negl Trop Dis. 2025 Jul 3;19(7):e0013181. doi: 10.1371/journal.pntd.0013181. eCollection 2025 Jul. PLoS Negl Trop Dis. 2025. PMID: 40609048 Free PMC article.
-
Viral intra-host evolutionary dynamics revealed via serial passage of Japanese encephalitis virus in vitro.Virus Evol. 2023 Mar 28;9(1):veac103. doi: 10.1093/ve/veac103. eCollection 2023. Virus Evol. 2023. PMID: 37205166 Free PMC article.
References
-
- Solomon T. Flavivirus encephalitis. The New England Journal of Medicine. 2004;351(4):370–409. - PubMed
-
- Lindenbach BD, Rice CM. Flaviviridae: the viruses and their replication. In: Knipe DM, Howley PM, editors. Fields Virology. 4th edition. Vol. 1. Philadelphia, Pa, USA: Lippincott Williams & Wilkins; 2001. pp. 991–1041.
-
- Sumiyoshi H, Mori C, Fuke I, et al. Complete nucleotide sequence of the Japanese encephalitis virus genome RNA. Virology. 1987;161(2):497–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
