

What controls DNA looping?
- PMID: 25167135
- PMCID: PMC4200792
- DOI: 10.3390/ijms150915090
What controls DNA looping?
Abstract
The looping of DNA provides a means of communication between sequentially distant genomic sites that operate in tandem to express, copy, and repair the information encoded in the DNA base sequence. The short loops implicated in the expression of bacterial genes suggest that molecular factors other than the naturally stiff double helix are involved in bringing the interacting sites into close spatial proximity. New computational techniques that take direct account of the three-dimensional structures and fluctuations of protein and DNA allow us to examine the likely means of enhancing such communication. Here, we describe the application of these approaches to the looping of a 92 base-pair DNA segment between the headpieces of the tetrameric Escherichia coli Lac repressor protein. The distortions of the double helix induced by a second protein--the nonspecific nucleoid protein HU--increase the computed likelihood of looping by several orders of magnitude over that of DNA alone. Large-scale deformations of the repressor, sequence-dependent features in the DNA loop, and deformability of the DNA operators also enhance looping, although to lesser degrees. The correspondence between the predicted looping propensities and the ease of looping derived from gene-expression and single-molecule measurements lends credence to the derived structural picture.
Figures
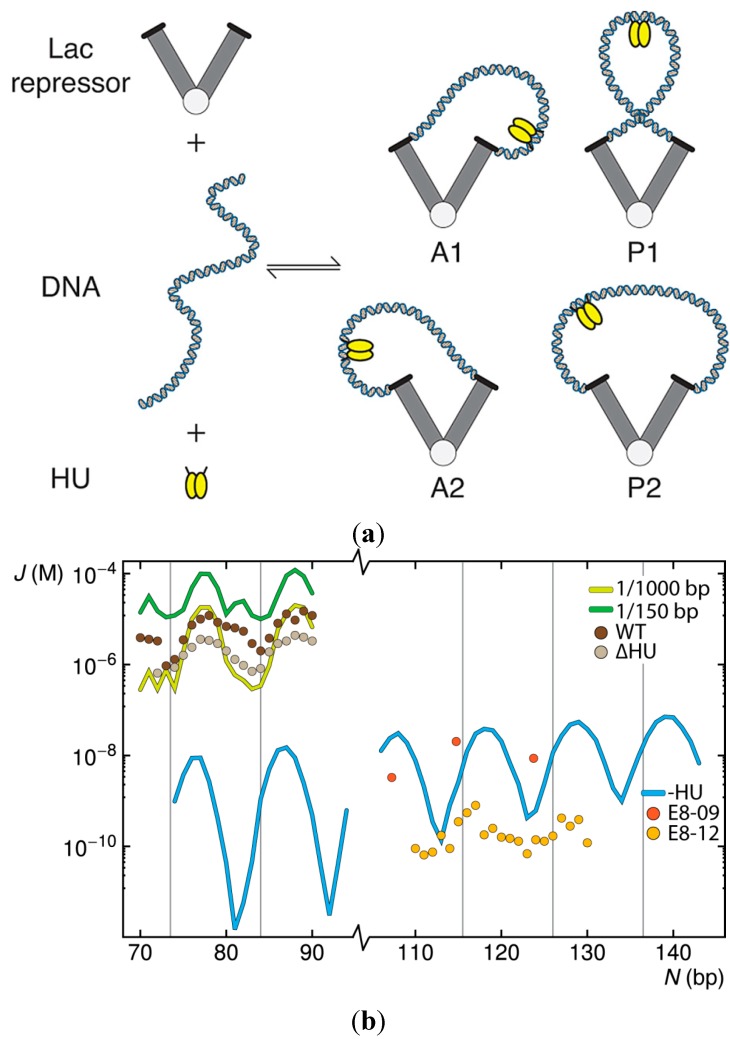
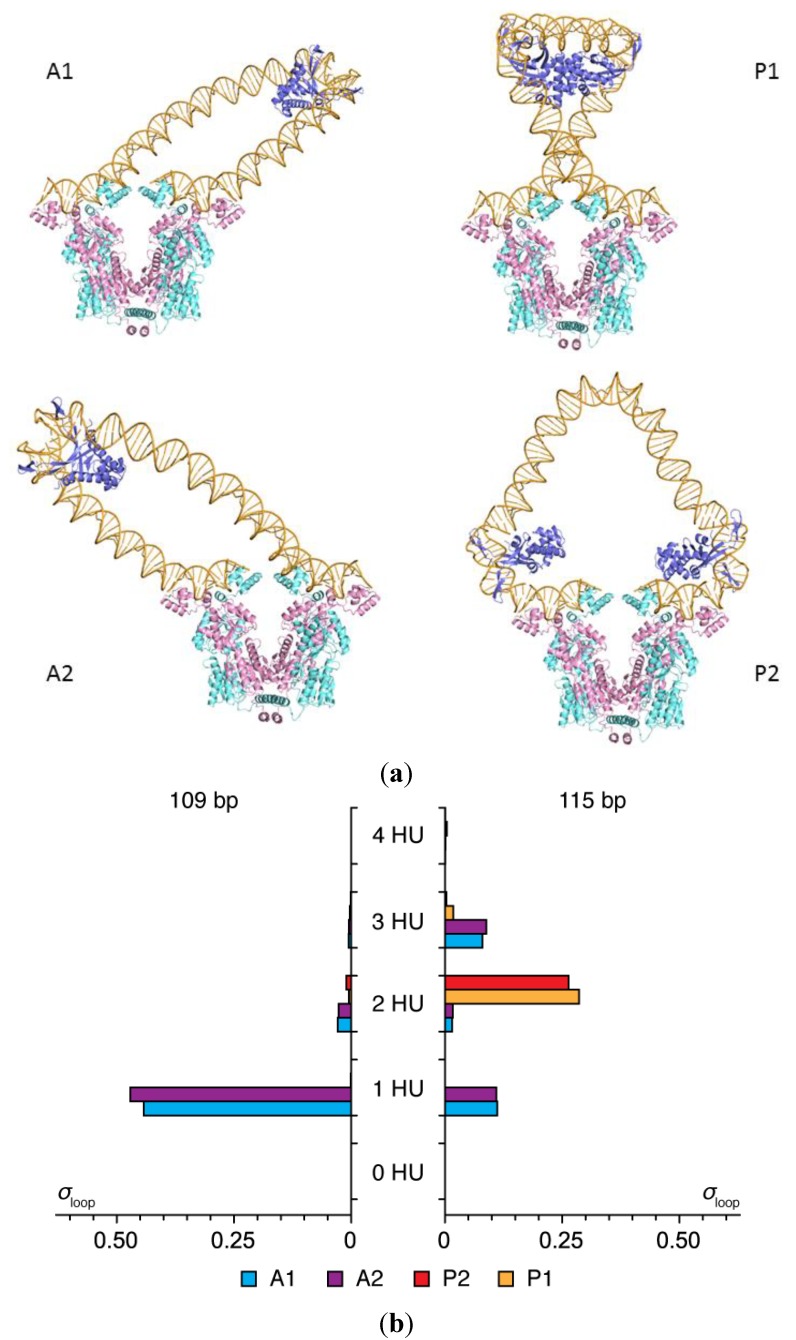
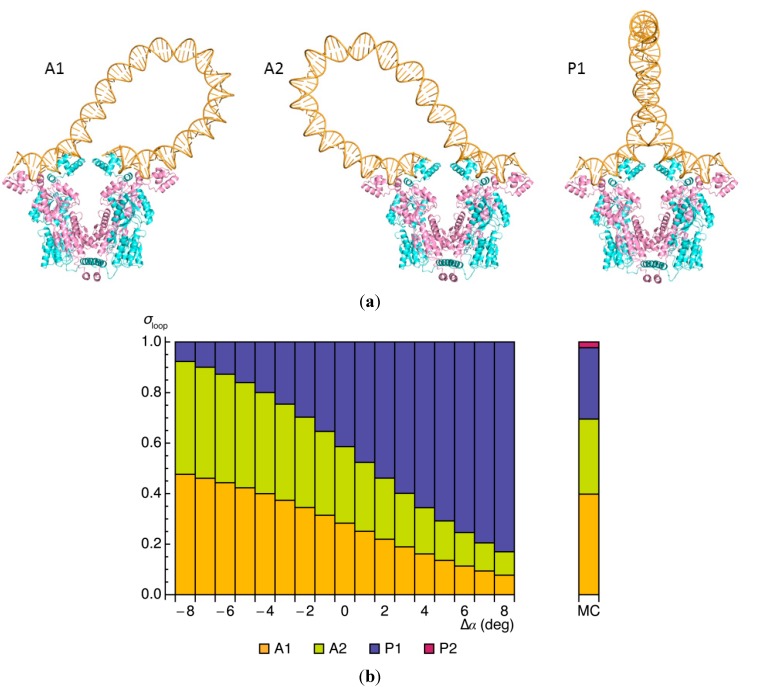
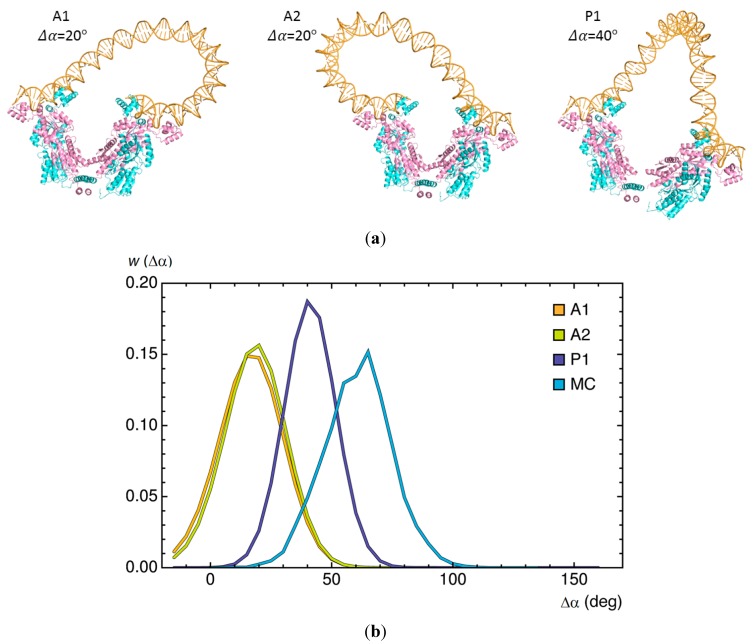
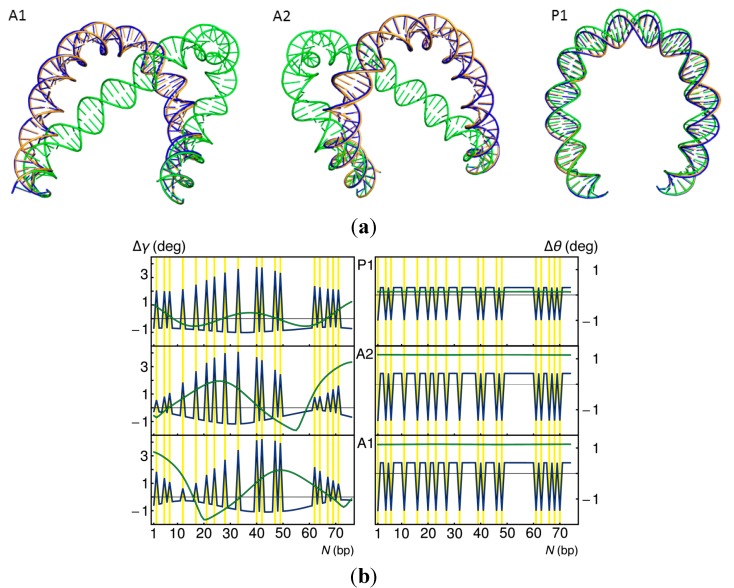
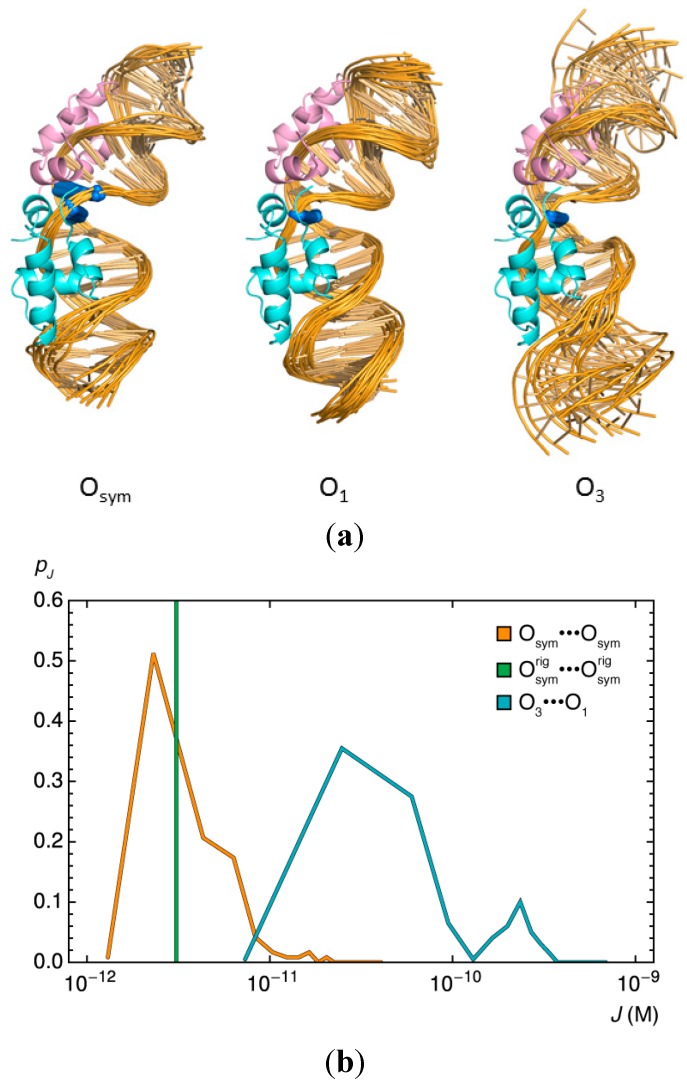
 ). Note the enhancement in looping with the natural operators.
). Note the enhancement in looping with the natural operators.References
-
- Müller-Hill B. The lac Operon. Walter de Gruyter; Berlin, Germany: 1996.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
