

Higher resources decrease fluctuating selection during host-parasite coevolution
- PMID: 25167763
- PMCID: PMC4257576
- DOI: 10.1111/ele.12337
Higher resources decrease fluctuating selection during host-parasite coevolution
Abstract
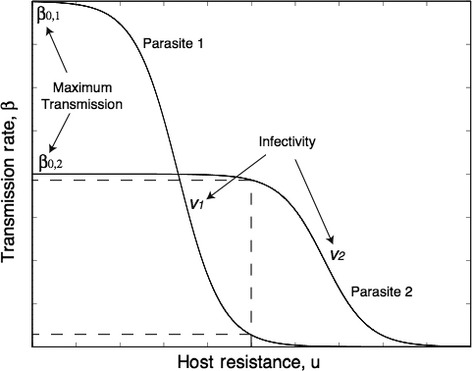
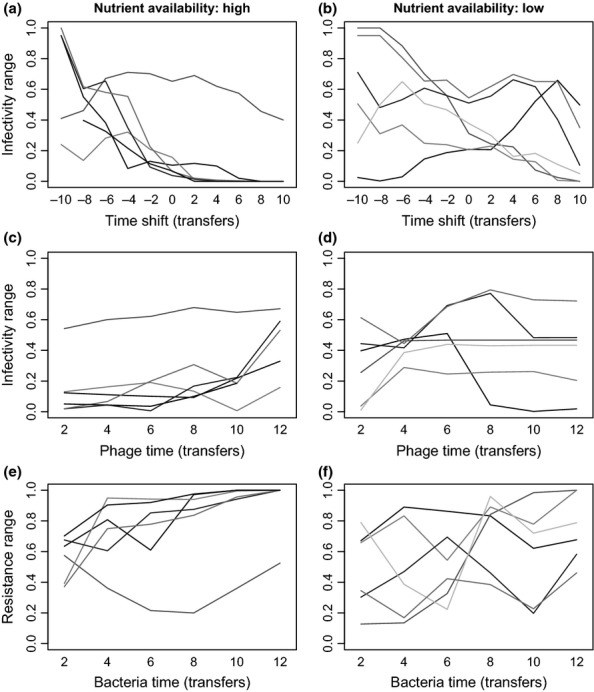
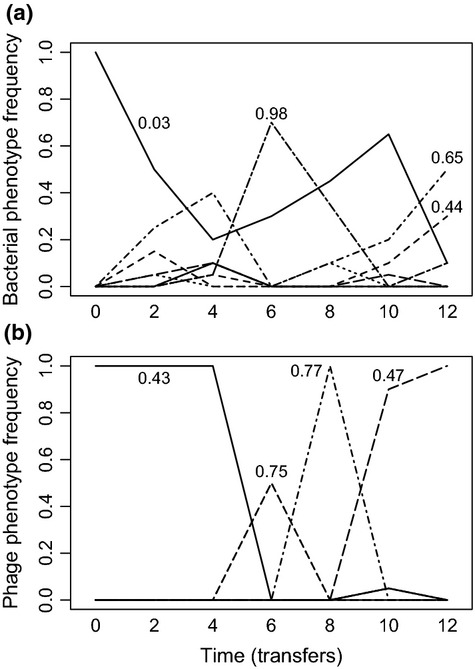
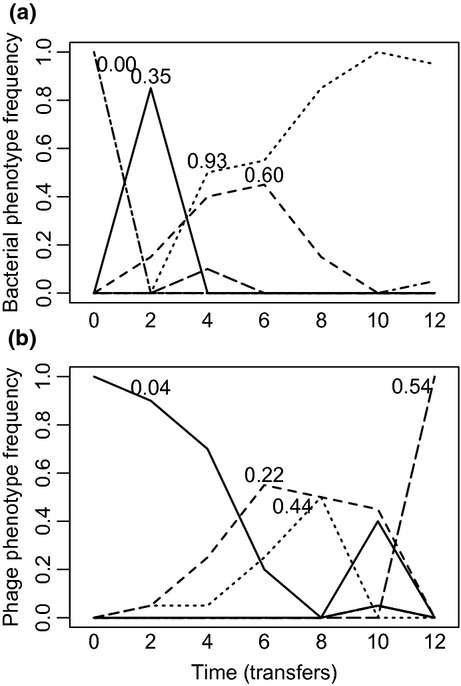
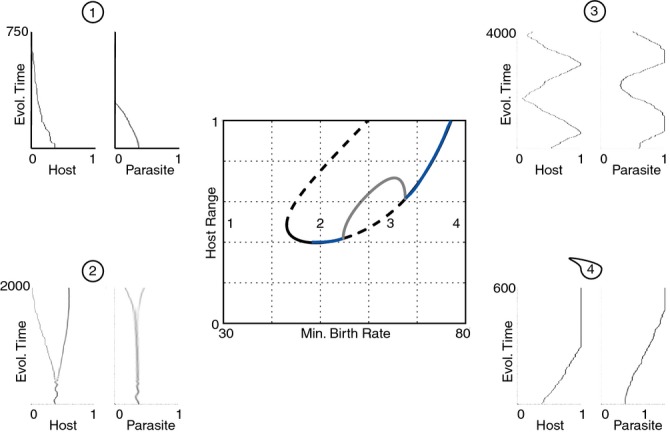
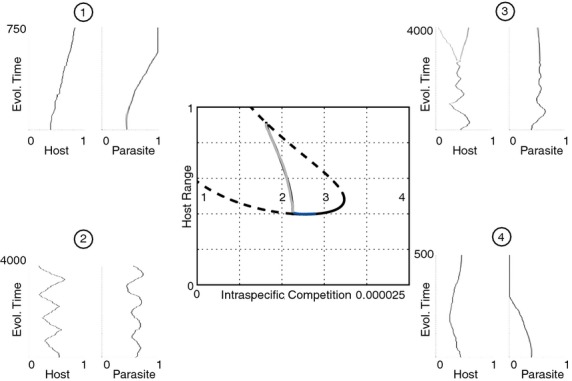
We still know very little about how the environment influences coevolutionary dynamics. Here, we investigated both theoretically and empirically how nutrient availability affects the relative extent of escalation of resistance and infectivity (arms race dynamic; ARD) and fluctuating selection (fluctuating selection dynamic; FSD) in experimentally coevolving populations of bacteria and viruses. By comparing interactions between clones of bacteria and viruses both within- and between-time points, we show that increasing nutrient availability resulted in coevolution shifting from FSD, with fluctuations in average infectivity and resistance ranges over time, to ARD. Our model shows that range fluctuations with lower nutrient availability can be explained both by elevated costs of resistance (a direct effect of nutrient availability), and reduced benefits of resistance when population sizes of hosts and parasites are lower (an indirect effect). Nutrient availability can therefore predictably and generally affect qualitative coevolutionary dynamics by both direct and indirect (mediated through ecological feedbacks) effects on costs of resistance.
Keywords: Adaptive dynamics; bacteria; experimental evolution; virus.
© 2014 The Authors. Ecology Letters published by John Wiley & Sons Ltd and CNRS.
Figures
References
-
- Agrawal A. Lively CM. Infection genetics: gene-for-gene versus matching-alleles models and all points in between. Evol. Ecol. Res. 2002;4:79–90.
-
- Anderson RM. May RM. The population dynamics of microparasites and their invertebrate hosts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1981;291:451–524.
-
- Anderson RM. May RM. Coevolution of hosts and parasites. Parasitology. 1982;85:411–426. - PubMed
-
- Bell G. The ecology and genetics of fitness in chlamydomonas.1. Genotype-by-environment interaction among pure strains. Proc. R. Soc. Lond. B. 1990;240:295–321.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
