

Evolution of resistance to a last-resort antibiotic in Staphylococcus aureus via bacterial competition
- PMID: 25171407
- PMCID: PMC4163622
- DOI: 10.1016/j.cell.2014.06.046
Evolution of resistance to a last-resort antibiotic in Staphylococcus aureus via bacterial competition
Abstract
Antibiotic resistance is a key medical concern, with antibiotic use likely being an important cause. However, here we describe an alternative route to clinically relevant antibiotic resistance that occurs solely due to competitive interactions among bacterial cells. We consistently observe that isolates of Methicillin-resistant Staphylococcus aureus diversify spontaneously into two distinct, sequentially arising strains. The first evolved strain outgrows the parent strain via secretion of surfactants and a toxic bacteriocin. The second is resistant to the bacteriocin. Importantly, this second strain is also resistant to intermediate levels of vancomycin. This so-called VISA (vancomycin-intermediate S. aureus) phenotype is seen in many hard-to-treat clinical isolates. This strain diversification also occurs during in vivo infection in a mouse model, which is consistent with the fact that both coevolved phenotypes resemble strains commonly found in clinic. Our study shows how competition between coevolving bacterial strains can generate antibiotic resistance and recapitulate key clinical phenotypes.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
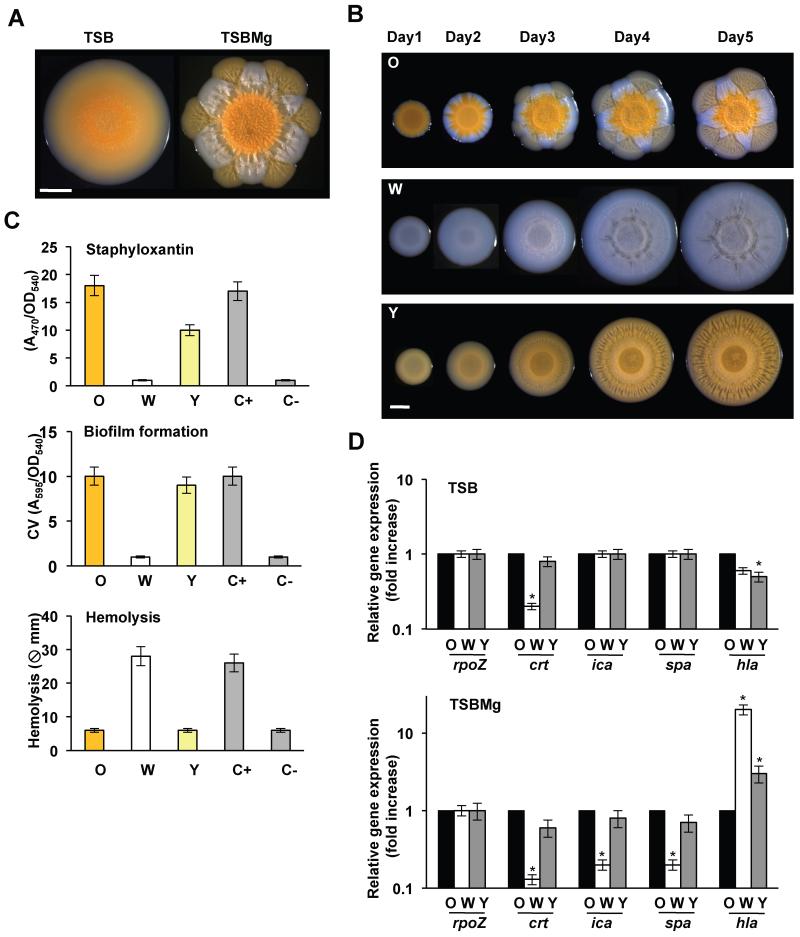
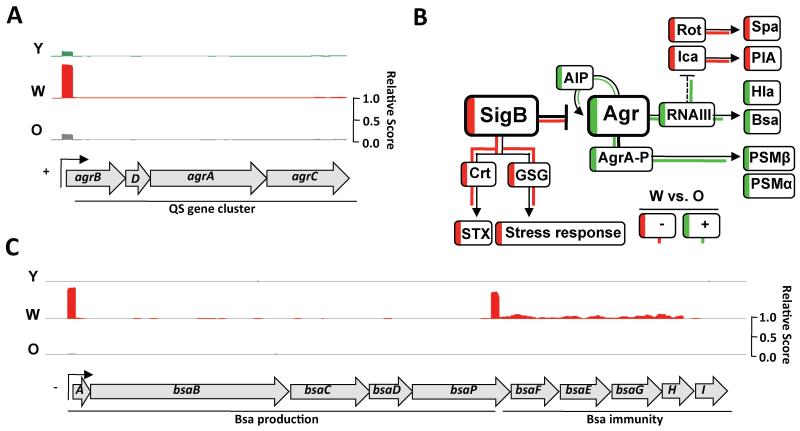
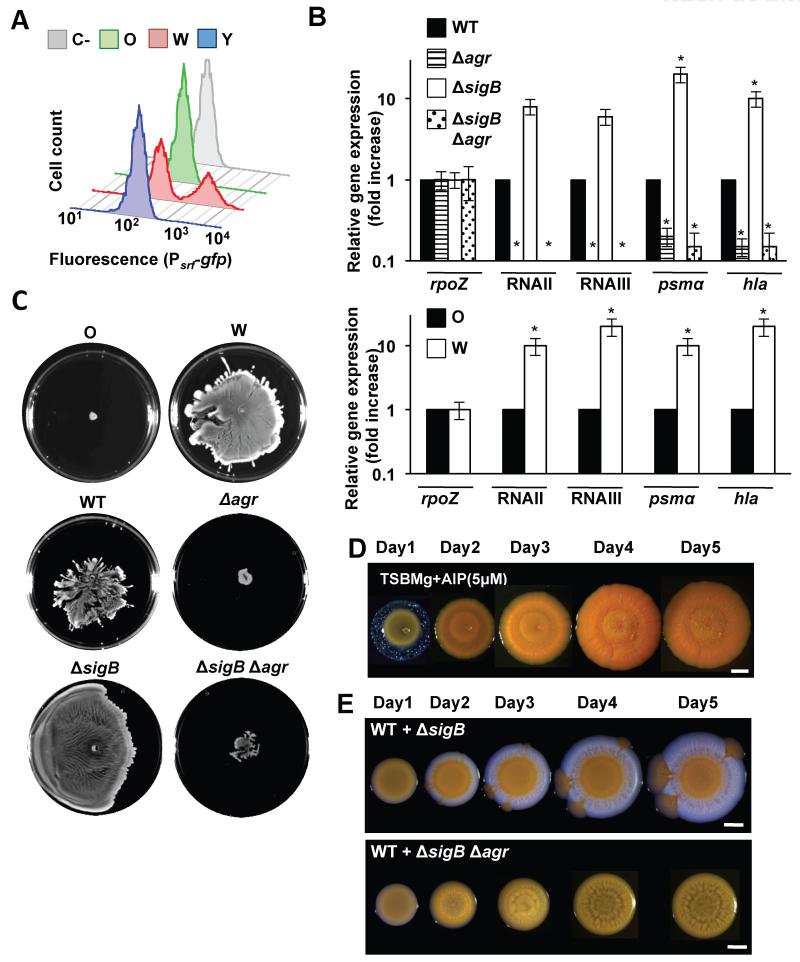
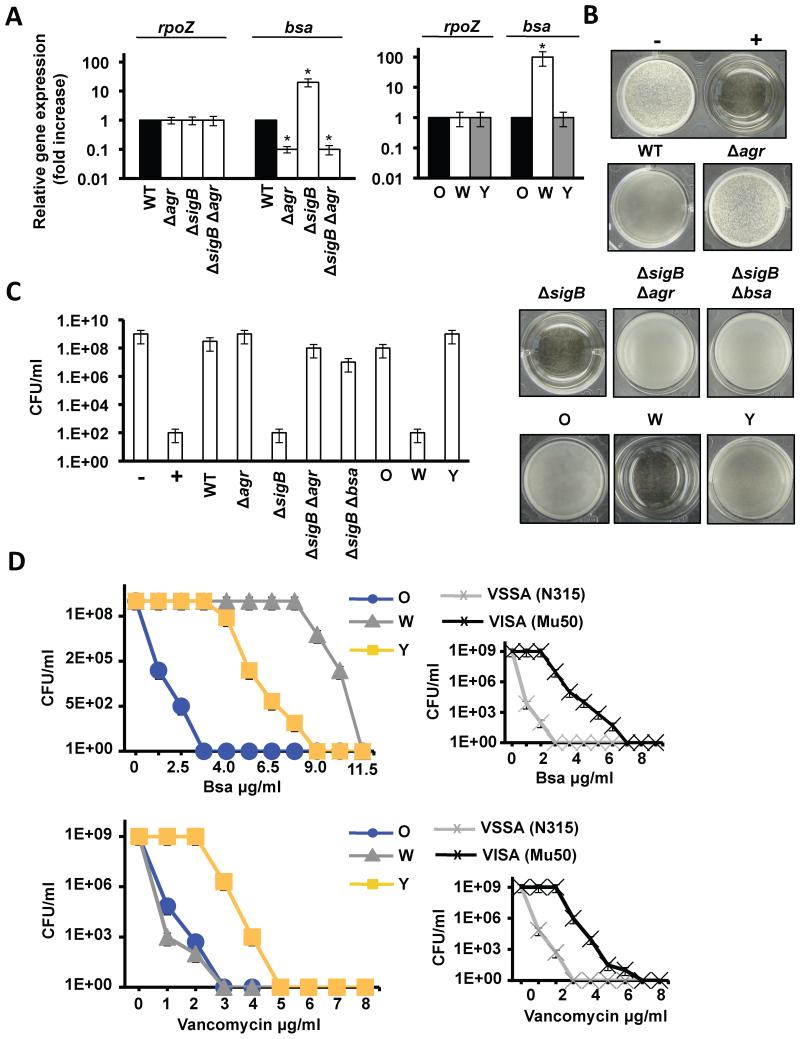
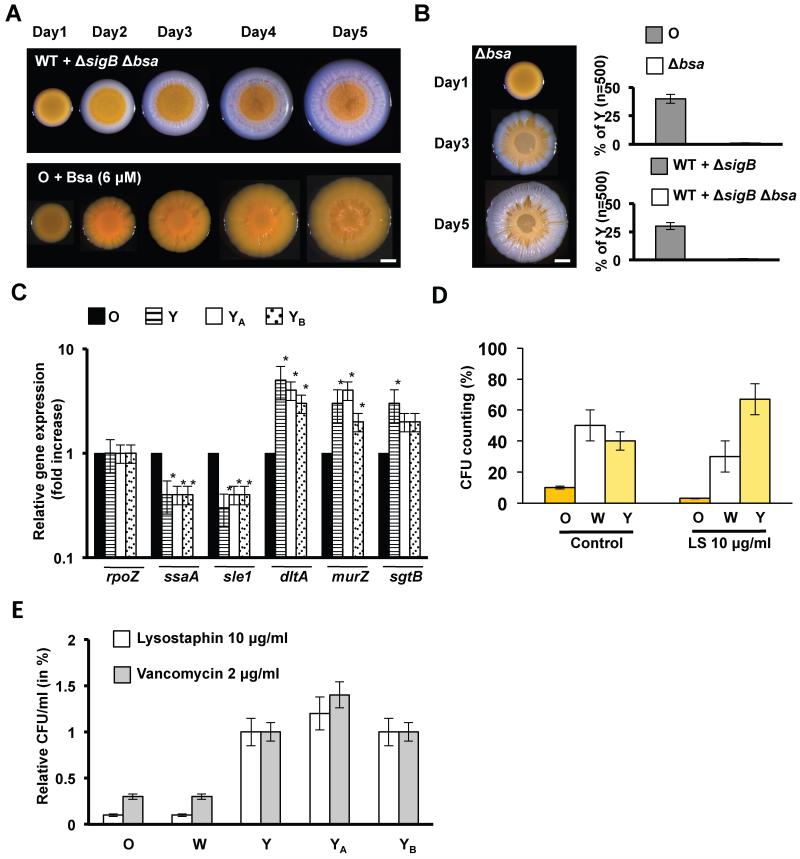
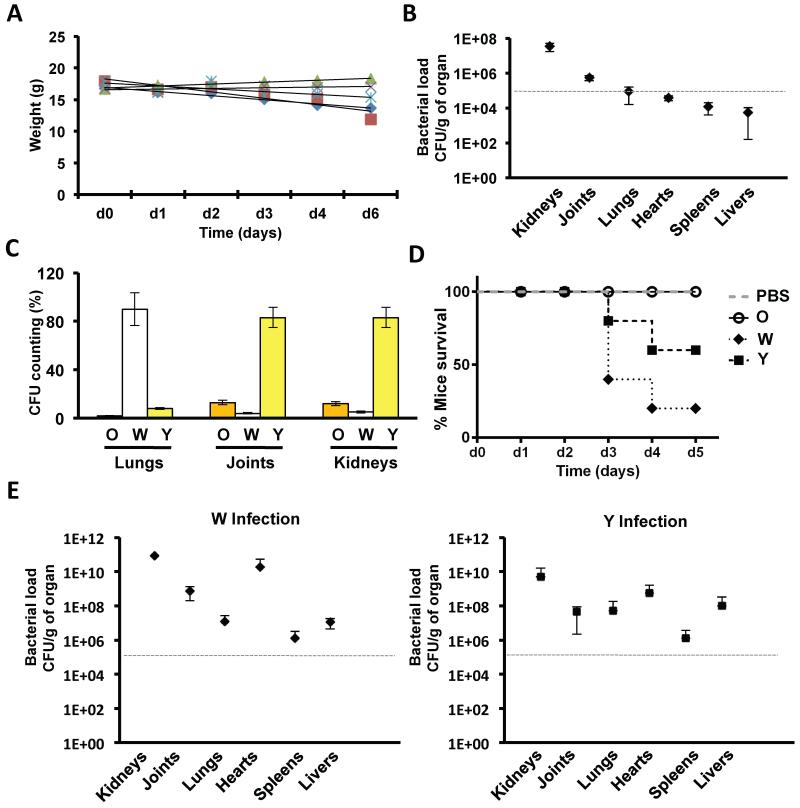
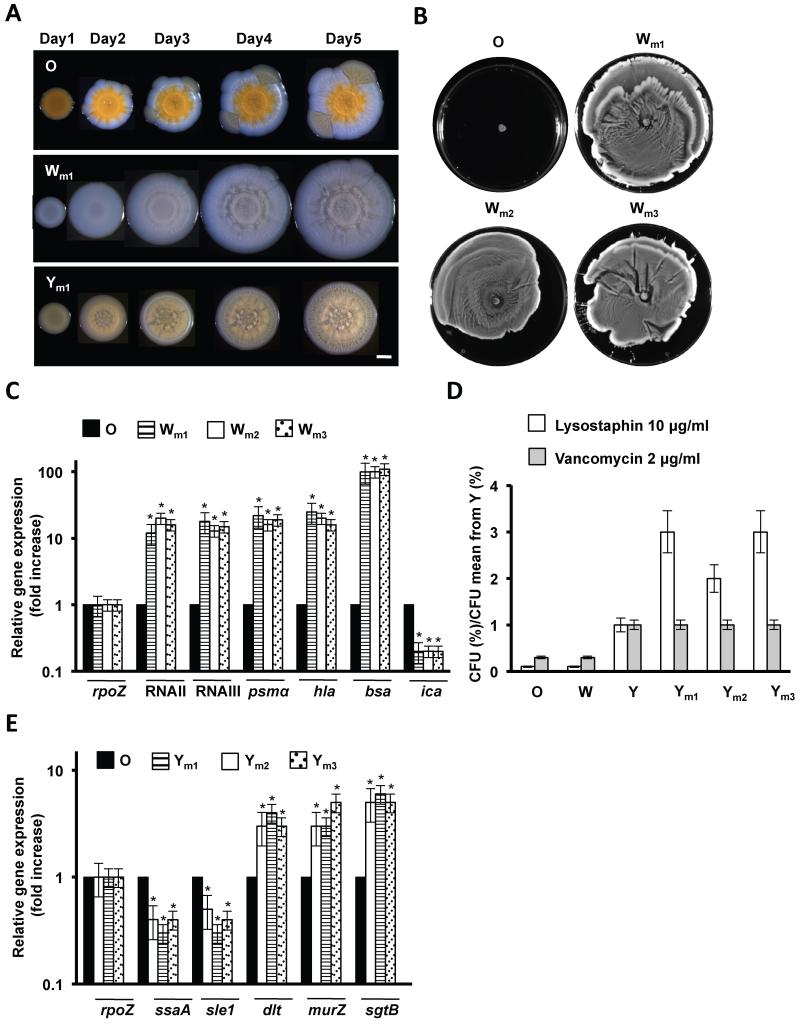
References
-
- Allen HK, Donato J, Wang HH, Cloud-Hansen KA, Davies J, Handelsman J. Call of the wild: antibiotic resistance genes in natural environments. Nature reviews Microbiology. 2010;8:251–259. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
