

An evolutionarily conserved prion-like element converts wild fungi from metabolic specialists to generalists
- PMID: 25171408
- PMCID: PMC4424049
- DOI: 10.1016/j.cell.2014.07.024
An evolutionarily conserved prion-like element converts wild fungi from metabolic specialists to generalists
Abstract
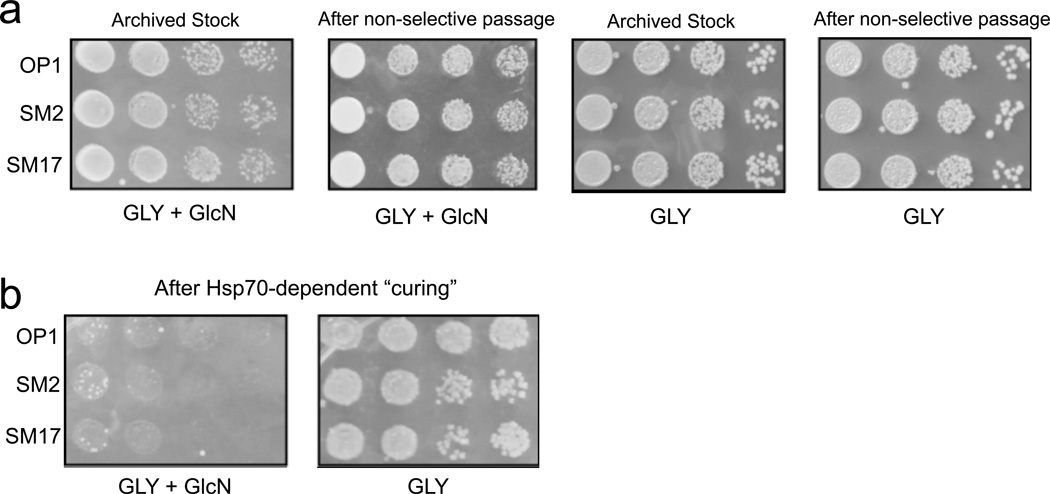
[GAR(+)] is a protein-based element of inheritance that allows yeast (Saccharomyces cerevisiae) to circumvent a hallmark of their biology: extreme metabolic specialization for glucose fermentation. When glucose is present, yeast will not use other carbon sources. [GAR(+)] allows cells to circumvent this "glucose repression." [GAR(+)] is induced in yeast by a factor secreted by bacteria inhabiting their environment. We report that de novo rates of [GAR(+)] appearance correlate with the yeast's ecological niche. Evolutionarily distant fungi possess similar epigenetic elements that are also induced by bacteria. As expected for a mechanism whose adaptive value originates from the selective pressures of life in biological communities, the ability of bacteria to induce [GAR(+)] and the ability of yeast to respond to bacterial signals have been extinguished repeatedly during the extended monoculture of domestication. Thus, [GAR(+)] is a broadly conserved adaptive strategy that links environmental and social cues to heritable changes in metabolism.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures






References
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting Proteostasis for Disease Intervention. Science. 2008;319:916–919. - PubMed
-
- Beaumont HJE, Gallie J, Kost C, Ferguson GC, Rainey PB. Experimental evolution of bet hedging. Nature. 2009;462:90–93. - PubMed
-
- Bisson LF, Karpel JE, Ramakrishnan V, Joseph L. Functional Genomics of Wine Yeast Saccharomyces cerevisiae. In: Taylor Steve L., editor. Advances in Food and Nutrition Research. Academic Press; 2007. pp. 65–121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
