

Axonally synthesized ATF4 transmits a neurodegenerative signal across brain regions
- PMID: 25171414
- PMCID: PMC4149755
- DOI: 10.1016/j.cell.2014.07.001
Axonally synthesized ATF4 transmits a neurodegenerative signal across brain regions
Abstract
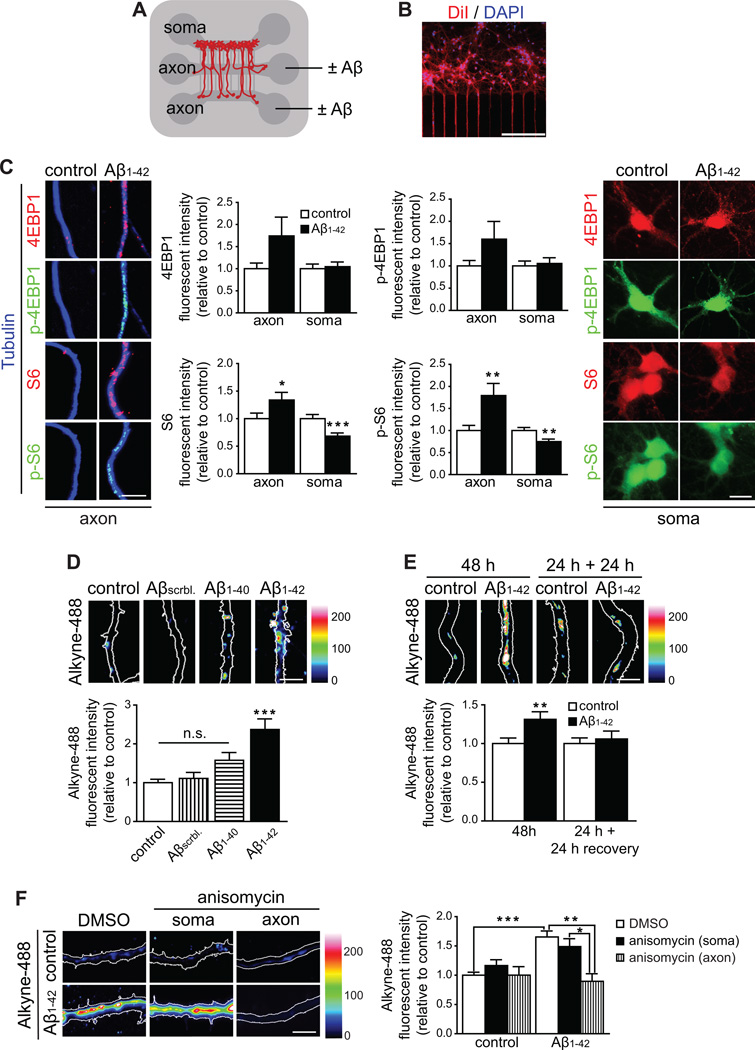
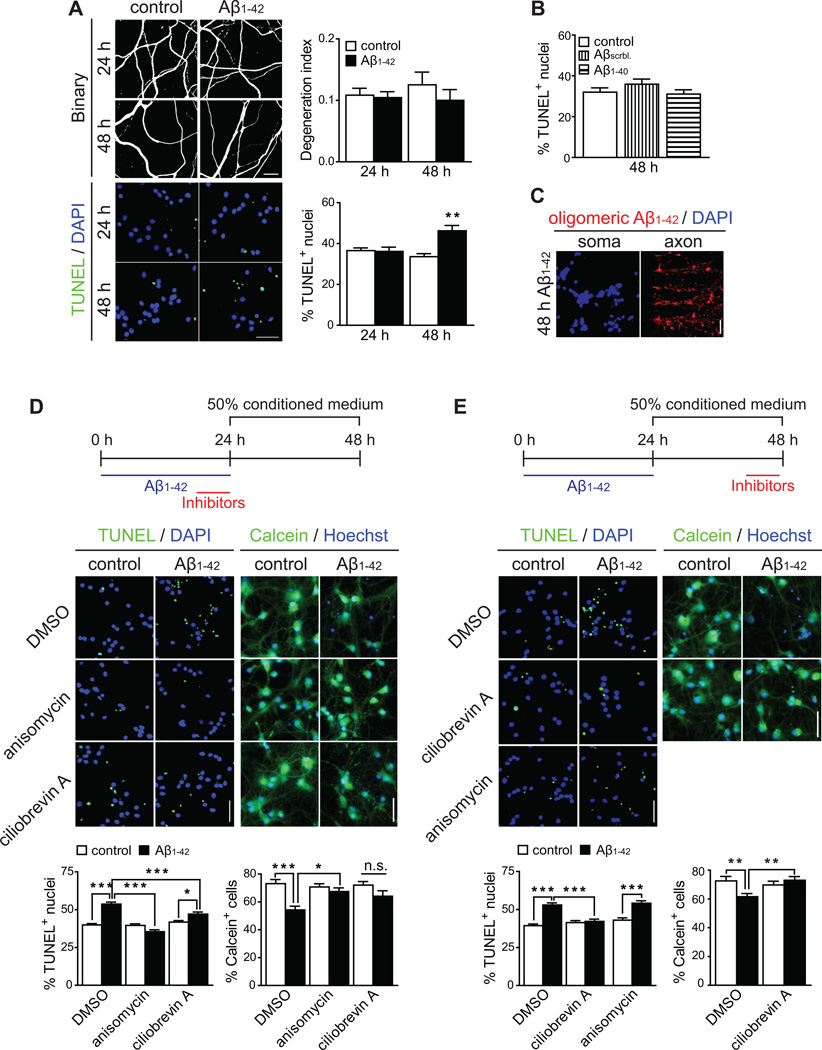
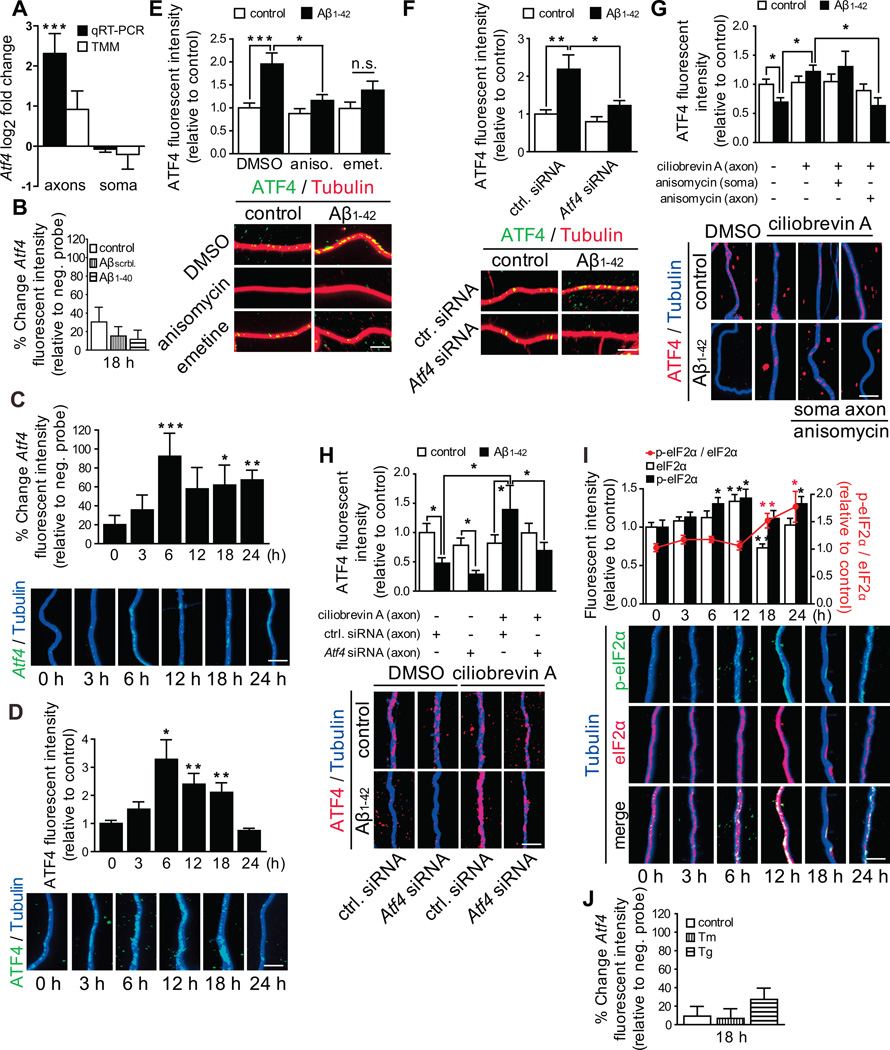
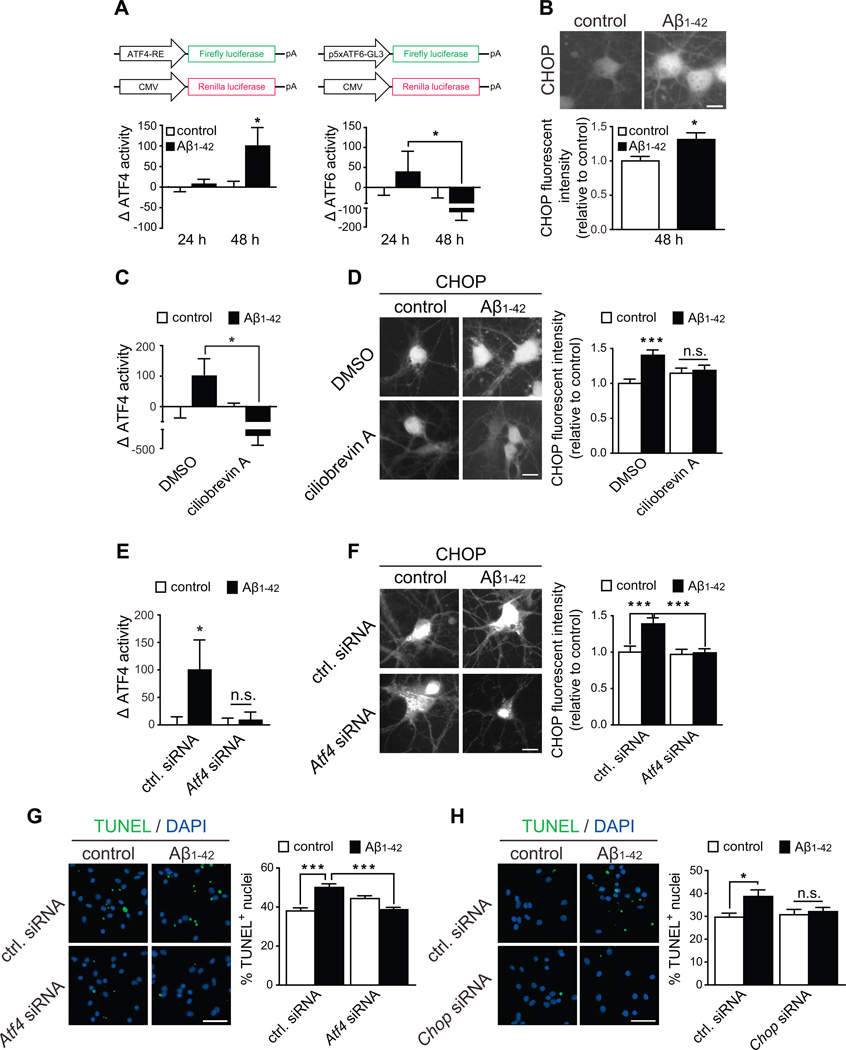
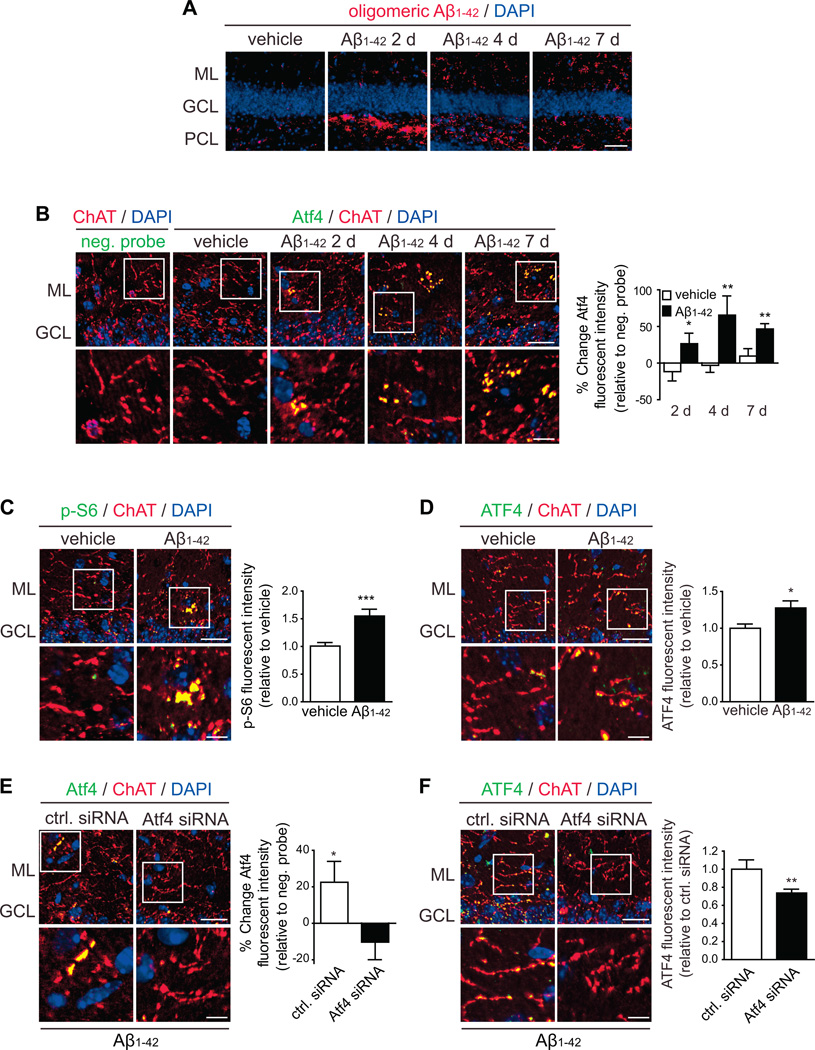
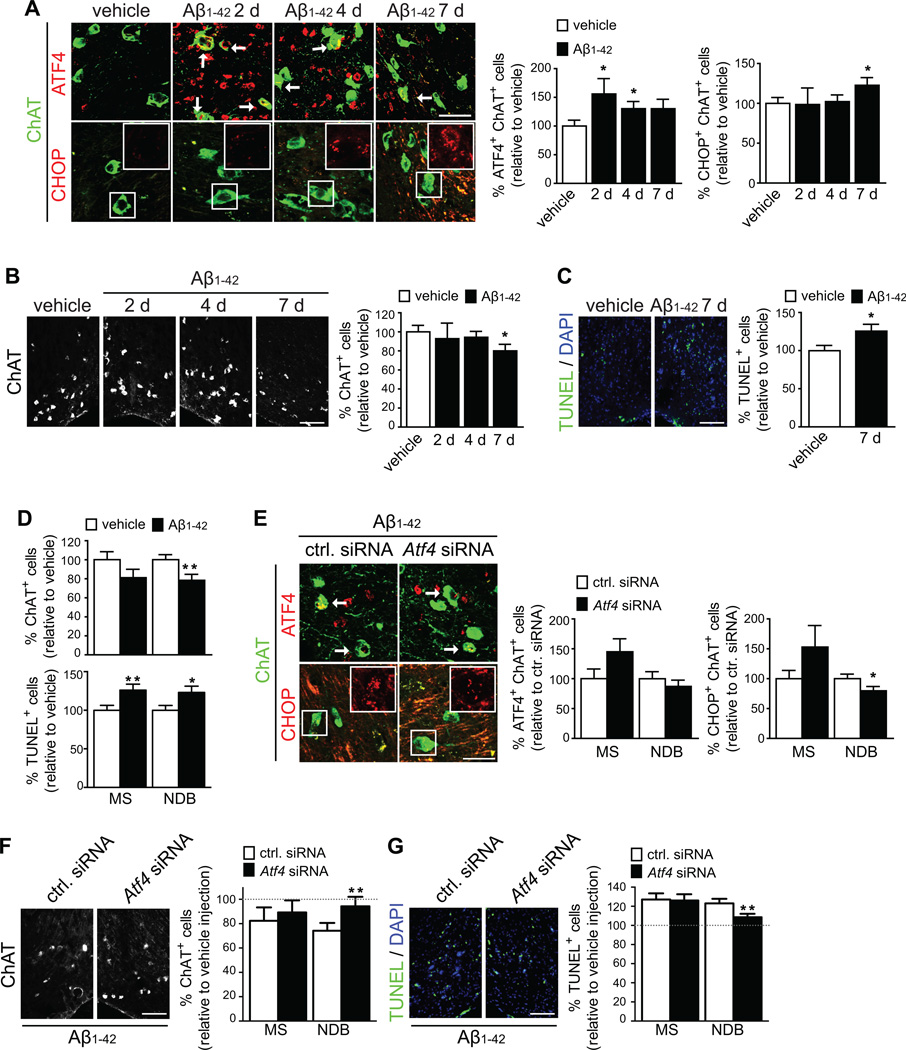
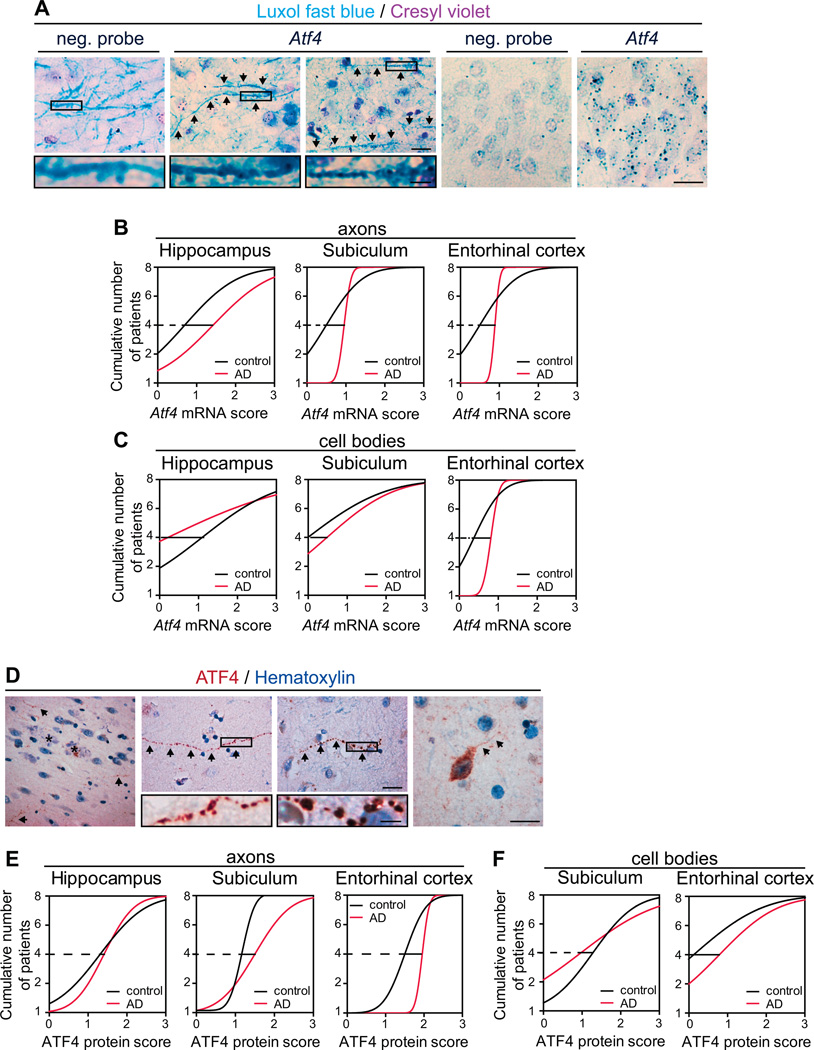
In Alzheimer's disease (AD) brain, exposure of axons to Aβ causes pathogenic changes that spread retrogradely by unknown mechanisms, affecting the entire neuron. We found that locally applied Aβ1-42 initiates axonal synthesis of a defined set of proteins including the transcription factor ATF4. Inhibition of local translation and retrograde transport or knockdown of axonal Atf4 mRNA abolished Aβ-induced ATF4 transcriptional activity and cell loss. Aβ1-42 injection into the dentate gyrus (DG) of mice caused loss of forebrain neurons whose axons project to the DG. Protein synthesis and Atf4 mRNA were upregulated in these axons, and coinjection of Atf4 siRNA into the DG reduced the effects of Aβ1-42 in the forebrain. ATF4 protein and transcripts were found with greater frequency in axons in the brain of AD patients. These results reveal an active role for intra-axonal translation in neurodegeneration and identify ATF4 as a mediator for the spread of AD pathology.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
ATF4: the perpetrator in axonal-mediated neurodegeneration in Alzheimer's disease.CNS Neurol Disord Drug Targets. 2014;13(9):1483-4. doi: 10.2174/1871527313666141110125829. CNS Neurol Disord Drug Targets. 2014. PMID: 25382233 No abstract available.
References
-
- Ameri K, Harris AL. Activating transcription factor 4. Int J Biochem Cell Biol. 2008;40:14–21. - PubMed
-
- Averous J, Bruhat A, Jousse C, Carraro V, Thiel G, Fafournoux P. Induction of CHOP expression by amino acid limitation requires both ATF4 expression and ATF2 phosphorylation. J Biol Chem. 2004;279:5288–5297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
