

Architecture of tight junctions and principles of molecular composition
- PMID: 25171873
- PMCID: PMC4254347
- DOI: 10.1016/j.semcdb.2014.08.011
Architecture of tight junctions and principles of molecular composition
Abstract
The tight junction creates an intercellular barrier limiting paracellular movement of solutes and material across epithelia. Currently many proteins have been identified as components of the tight junction and understanding their architectural organization and interactions is critical to understanding the biology of the barrier. In general the architecture can be conceptualized into compartments with the transmembrane barrier proteins (claudins, occludin, JAM-A, etc.), linked to peripheral scaffolding proteins (such as ZO-1, afadin, MAGI1, etc.) which are in turned linked to actin and microtubules through numerous linkers (cingulin, myosins, protein 4.1, etc.). Within this complex network are associated many signaling proteins that affect the barrier and broader cell functions. The PDZ domain is a commonly used motif to specifically link individual junction protein pairs. Here we review some of the key proteins defining the tight junction and general themes of their organization with the perspective that much will be learned about function by characterizing the detailed architecture and subcompartments within the junction.
Keywords: Actin; Claudin; Epithelium; Occludin; Tight junction; ZO-1.
Published by Elsevier Ltd.
Figures
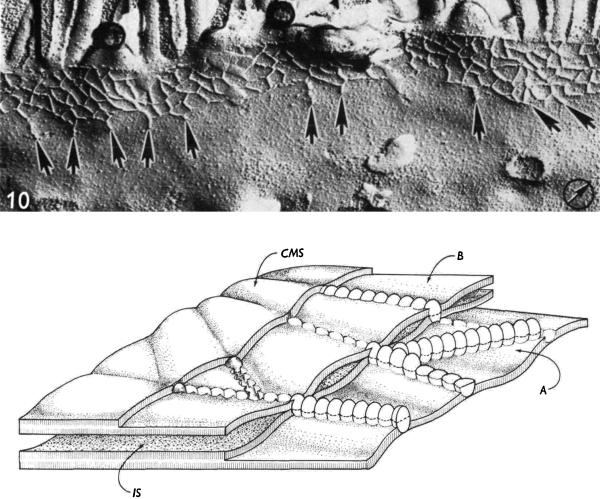
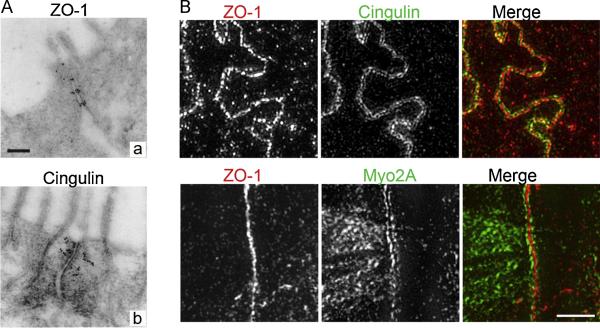
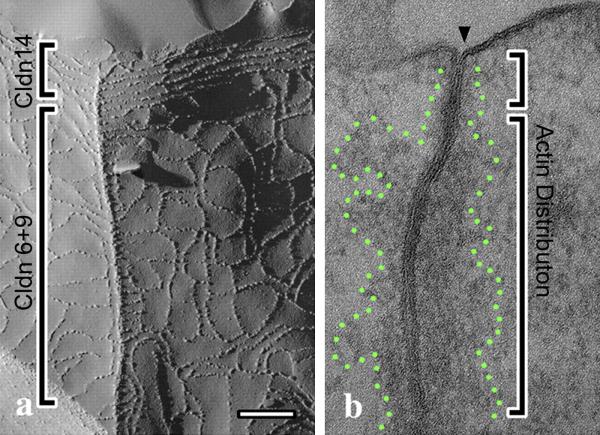
References
-
- Fromter E, Diamond J. Route of passive ion permeation in epithelia. Nat New Biol. 1972;235:9–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
