

Hybrid incompatibility arises in a sequence-based bioenergetic model of transcription factor binding
- PMID: 25173845
- PMCID: PMC4224158
- DOI: 10.1534/genetics.114.168112
Hybrid incompatibility arises in a sequence-based bioenergetic model of transcription factor binding
Abstract
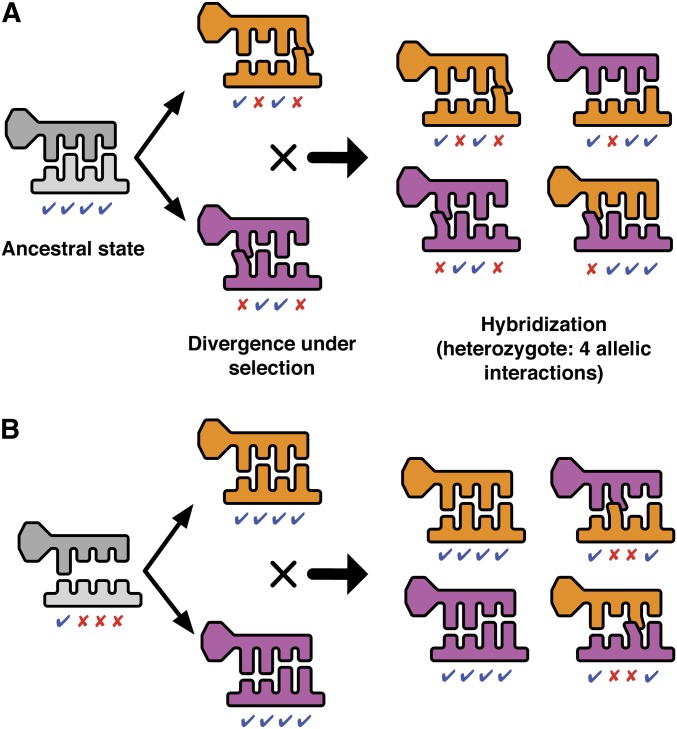
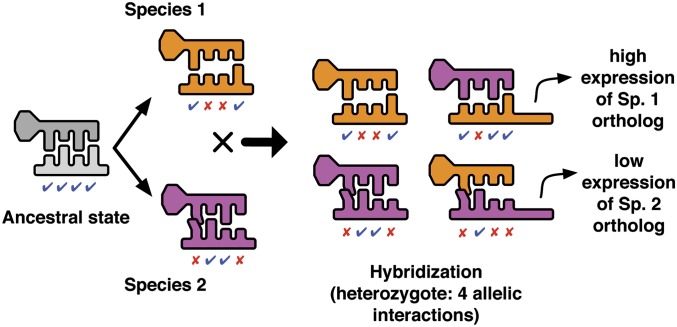
Postzygotic isolation between incipient species results from the accumulation of incompatibilities that arise as a consequence of genetic divergence. When phenotypes are determined by regulatory interactions, hybrid incompatibility can evolve even as a consequence of parallel adaptation in parental populations because interacting genes can produce the same phenotype through incompatible allelic combinations. We explore the evolutionary conditions that promote and constrain hybrid incompatibility in regulatory networks using a bioenergetic model (combining thermodynamics and kinetics) of transcriptional regulation, considering the bioenergetic basis of molecular interactions between transcription factors (TFs) and their binding sites. The bioenergetic parameters consider the free energy of formation of the bond between the TF and its binding site and the availability of TFs in the intracellular environment. Together these determine fractional occupancy of the TF on the promoter site, the degree of subsequent gene expression and in diploids, and the degree of dominance among allelic interactions. This results in a sigmoid genotype-phenotype map and fitness landscape, with the details of the shape determining the degree of bioenergetic evolutionary constraint on hybrid incompatibility. Using individual-based simulations, we subjected two allopatric populations to parallel directional or stabilizing selection. Misregulation of hybrid gene expression occurred under either type of selection, although it evolved faster under directional selection. Under directional selection, the extent of hybrid incompatibility increased with the slope of the genotype-phenotype map near the derived parental expression level. Under stabilizing selection, hybrid incompatibility arose from compensatory mutations and was greater when the bioenergetic properties of the interaction caused the space of nearly neutral genotypes around the stable expression level to be wide. F2's showed higher hybrid incompatibility than F1's to the extent that the bioenergetic properties favored dominant regulatory interactions. The present model is a mechanistically explicit case of the Bateson-Dobzhansky-Muller model, connecting environmental selective pressure to hybrid incompatibility through the molecular mechanism of regulatory divergence. The bioenergetic parameters that determine expression represent measurable properties of transcriptional regulation, providing a predictive framework for empirical studies of how phenotypic evolution results in epistatic incompatibility at the molecular level in hybrids.
Keywords: Dobzhansky–Muller interactions; adaptive landscape; cis–trans coevolution; genotype–phenotype map; regulatory evolution; speciation.
Copyright © 2014 by the Genetics Society of America.
Figures
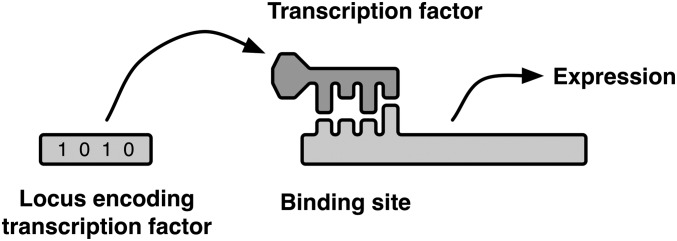
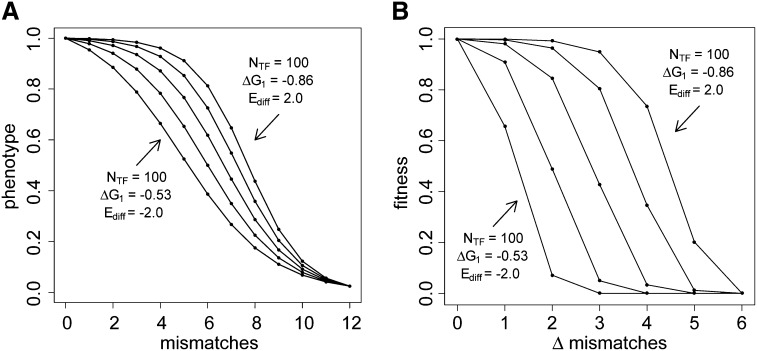
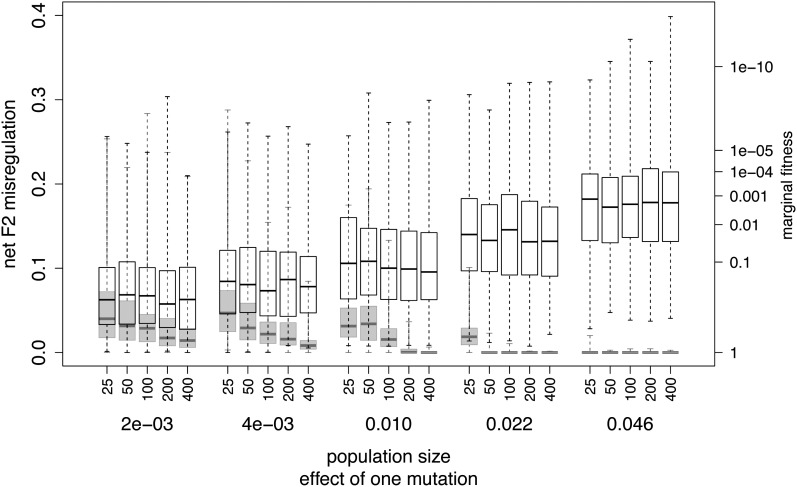
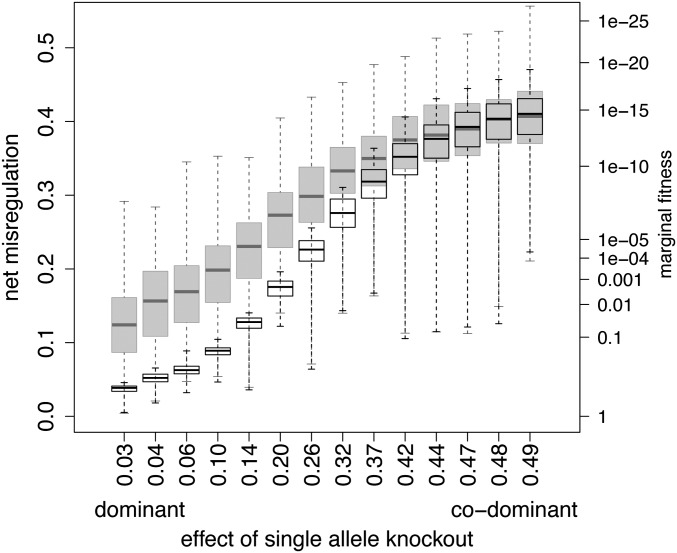
Similar articles
-
Hybrid incompatibility despite pleiotropic constraint in a sequence-based bioenergetic model of transcription factor binding.Genetics. 2014 Dec;198(4):1645-54. doi: 10.1534/genetics.114.171397. Epub 2014 Oct 13. Genetics. 2014. PMID: 25313130 Free PMC article.
-
Genetic architecture and postzygotic reproductive isolation: evolution of Bateson-Dobzhansky-Muller incompatibilities in a polygenic model.Evolution. 2010 Mar 1;64(3):675-93. doi: 10.1111/j.1558-5646.2009.00861.x. Epub 2009 Oct 5. Evolution. 2010. PMID: 19817852
-
Simple Biophysical Model Predicts Faster Accumulation of Hybrid Incompatibilities in Small Populations Under Stabilizing Selection.Genetics. 2015 Dec;201(4):1525-37. doi: 10.1534/genetics.115.181685. Epub 2015 Oct 4. Genetics. 2015. PMID: 26434721 Free PMC article.
-
The Ecology of Hybrid Incompatibilities.Cold Spring Harb Perspect Biol. 2024 Sep 3;16(9):a041440. doi: 10.1101/cshperspect.a041440. Cold Spring Harb Perspect Biol. 2024. PMID: 38151331 Review.
-
Evolution and Molecular Control of Hybrid Incompatibility in Plants.Front Plant Sci. 2016 Aug 11;7:1208. doi: 10.3389/fpls.2016.01208. eCollection 2016. Front Plant Sci. 2016. PMID: 27563306 Free PMC article. Review.
Cited by
-
The Pace of Hybrid Incompatibility Evolution in House Mice.Genetics. 2015 Sep;201(1):229-42. doi: 10.1534/genetics.115.179499. Epub 2015 Jul 20. Genetics. 2015. PMID: 26199234 Free PMC article.
-
Ecological divergence in sympatry causes gene misexpression in hybrids.Mol Ecol. 2020 Jul;29(14):2707-2721. doi: 10.1111/mec.15512. Epub 2020 Jul 10. Mol Ecol. 2020. PMID: 32557903 Free PMC article.
-
A New Mechanism for Mendelian Dominance in Regulatory Genetic Pathways: Competitive Binding by Transcription Factors.Genetics. 2017 Jan;205(1):101-112. doi: 10.1534/genetics.116.195255. Epub 2016 Nov 18. Genetics. 2017. PMID: 27866169 Free PMC article.
-
Gene Regulation and Speciation.Trends Genet. 2017 Jan;33(1):68-80. doi: 10.1016/j.tig.2016.11.003. Epub 2016 Dec 1. Trends Genet. 2017. PMID: 27914620 Free PMC article. Review.
-
Unifying framework explaining how parental regulatory divergence can drive gene expression in hybrids and allopolyploids.Nat Commun. 2024 Oct 8;15(1):8714. doi: 10.1038/s41467-024-52546-5. Nat Commun. 2024. PMID: 39379366 Free PMC article.
References
-
- Bateson W., 1909. Heredity and variation in modern lights, Darwin and Modern Science, edited by Seward A. C. Cambridge University Press, Cambridge, UK.
-
- Coyne J. A., Orr H. A., 2004. Speciation. Sinauer, Sunderland, MA.
-
- Dobzhansky T., 1937. Genetics and the Origin of Species. Columbia University Press, New York.
-
- Fierst J. L., Hansen T. F., 2010. Genetic architecture and postzygotic reproductive isolation: evolution of Bateson–Dobzhansky–Muller incompatibilities in a polygenic model. Evolution 64: 675–693. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
