

Dominant lethal pathologies in male mice engineered to contain an X-linked DUX4 transgene
- PMID: 25176645
- PMCID: PMC4188423
- DOI: 10.1016/j.celrep.2014.07.056
Dominant lethal pathologies in male mice engineered to contain an X-linked DUX4 transgene
Abstract
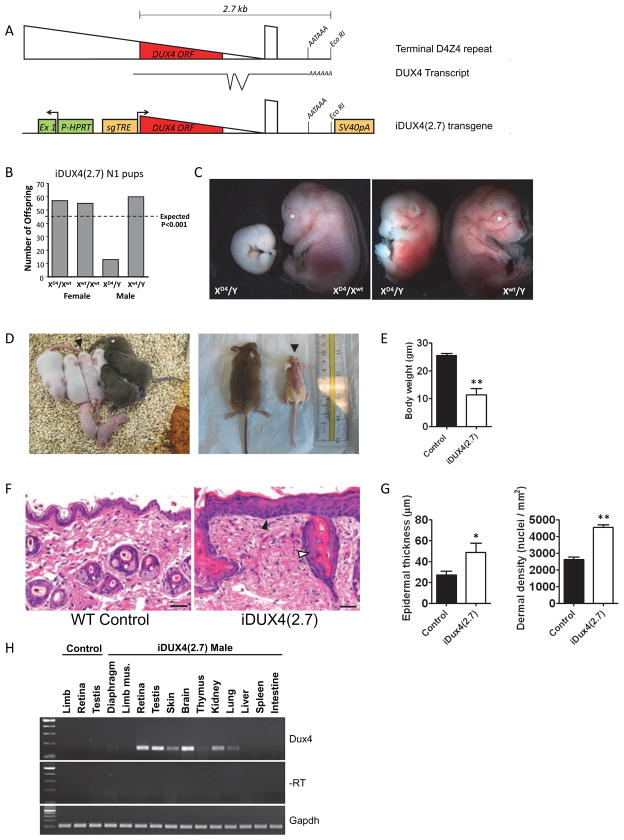
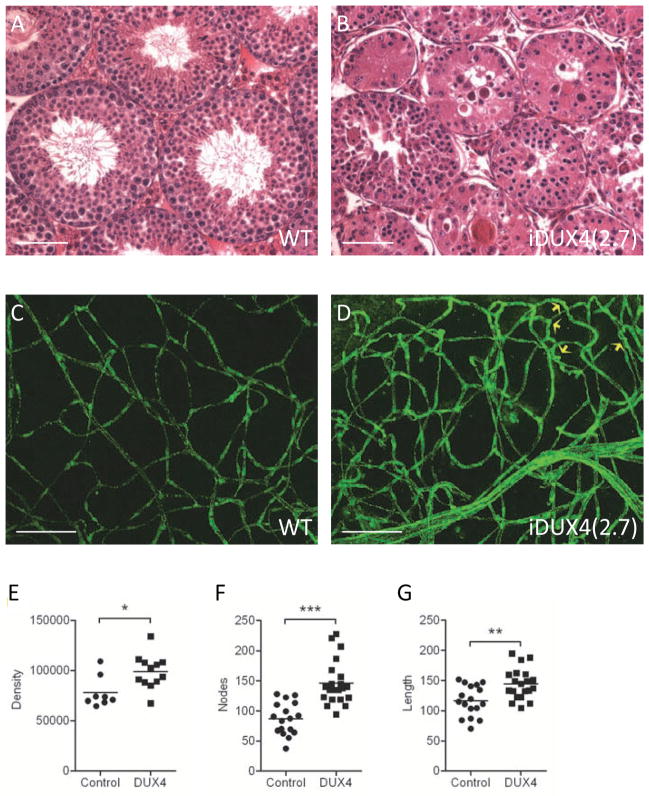
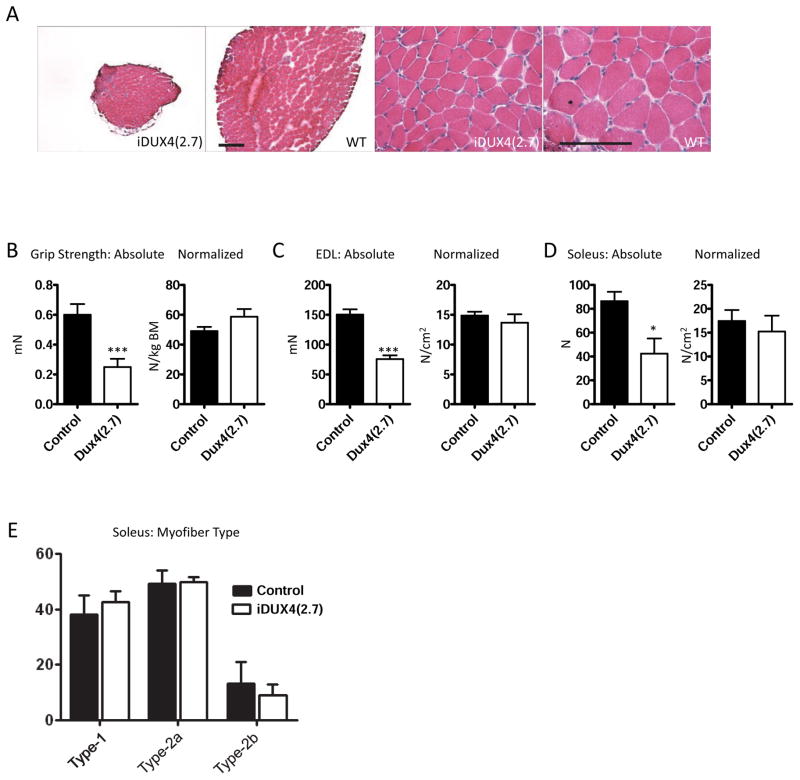
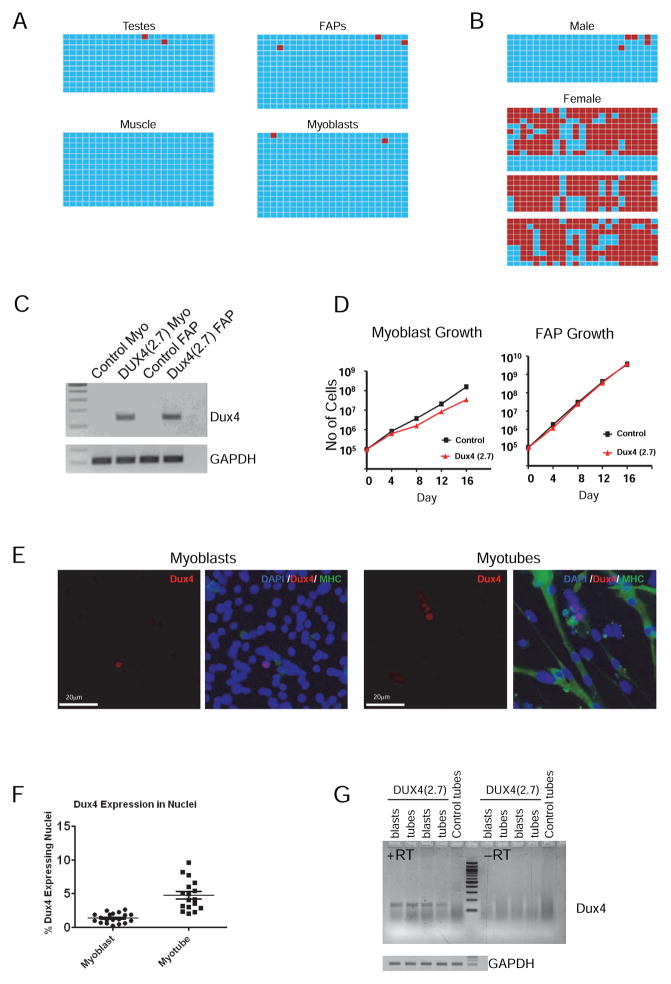
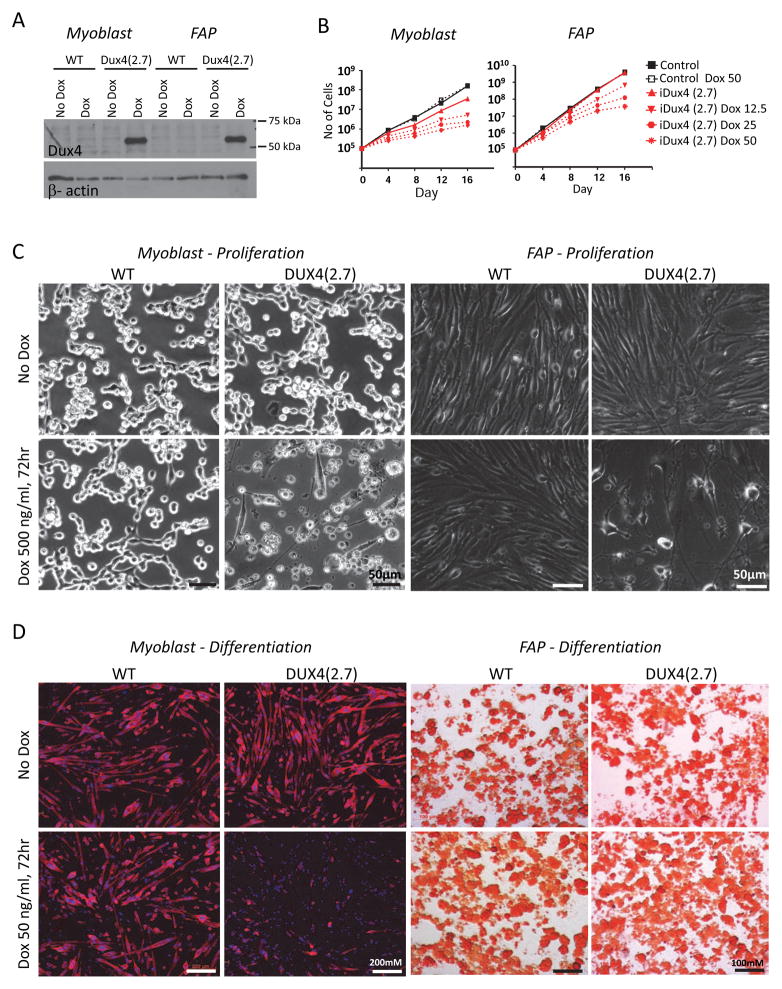
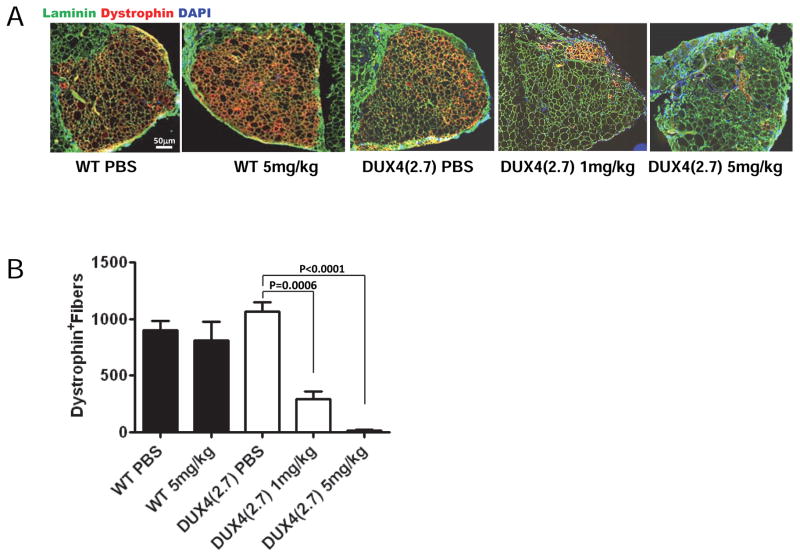
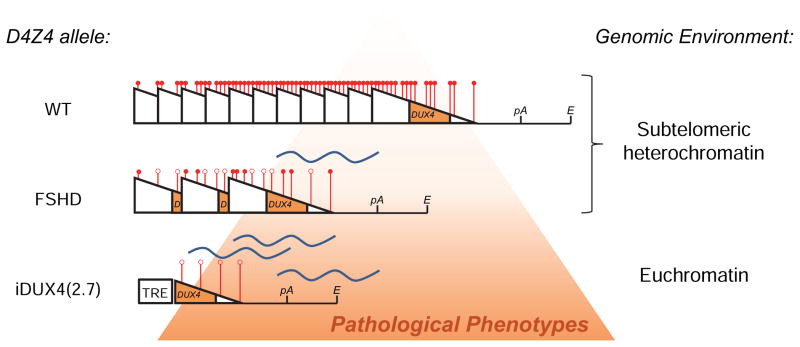
Facioscapulohumeral muscular dystrophy (FSHD) is an enigmatic disease associated with epigenetic alterations in the subtelomeric heterochromatin of the D4Z4 macrosatellite repeat. Each repeat unit encodes DUX4, a gene that is normally silent in most tissues. Besides muscular loss, most patients suffer retinal vascular telangiectasias. To generate an animal model, we introduced a doxycycline-inducible transgene encoding DUX4 and 3' genomic DNA into a euchromatic region of the mouse X chromosome. Without induction, DUX4 RNA was expressed at low levels in many tissues and animals displayed a variety of unexpected dominant leaky phenotypes, including male-specific lethality. Remarkably, rare live-born males expressed DUX4 RNA in the retina and presented a retinal vascular telangiectasia. By using doxycycline to induce DUX4 expression in satellite cells, we observed impaired myogenesis in vitro and in vivo. This mouse model, which shows pathologies due to FSHD-related D4Z4 sequences, is likely to be useful for testing anti-DUX4 therapies in FSHD.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Agha-Mohammadi S, O’Malley M, Etemad A, Wang Z, Xiao X, Lotze MT. Second-generation tetracycline-regulatable promoter: repositioned tet operator elements optimize transactivator synergy while shorter minimal promoter offers tight basal leakiness. J Gene Med. 2004;6:817–828. - PubMed
-
- Bakker E, Wijmenga C, Vossen RH, Padberg GW, Hewitt J, van der Wielen M, Rasmussen K, Frants RR. The FSHD-linked locus D4F104S1 (p13E-11) on 4q35 has a homologue on 10qter. Muscle Nerve. 1995;2:S39–44. - PubMed
-
- Bosnakovski D, Lamb S, Simsek T, Xu Z, Belayew A, Perlingeiro R, Kyba M. DUX4c, an FSHD candidate gene, interferes with myogenic regulators and abolishes myoblast differentiation. Exp Neurol 2008a - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL68802/HL/NHLBI NIH HHS/United States
- T32 AR007612/AR/NIAMS NIH HHS/United States
- R01 HD053889/HD/NICHD NIH HHS/United States
- R01 AR055685/AR/NIAMS NIH HHS/United States
- R01 HL103733/HL/NHLBI NIH HHS/United States
- P30 AR0507220/AR/NIAMS NIH HHS/United States
- K02-AG036827/AG/NIA NIH HHS/United States
- R01 HL103773/HL/NHLBI NIH HHS/United States
- R01 AR055299/AR/NIAMS NIH HHS/United States
- R01 HL068802/HL/NHLBI NIH HHS/United States
- K02 AG036827/AG/NIA NIH HHS/United States
- U01 HL100407/HL/NHLBI NIH HHS/United States
- R01AR055299/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
