

Neural correlates of short-term memory in primate auditory cortex
- PMID: 25177266
- PMCID: PMC4132374
- DOI: 10.3389/fnins.2014.00250
Neural correlates of short-term memory in primate auditory cortex
Abstract
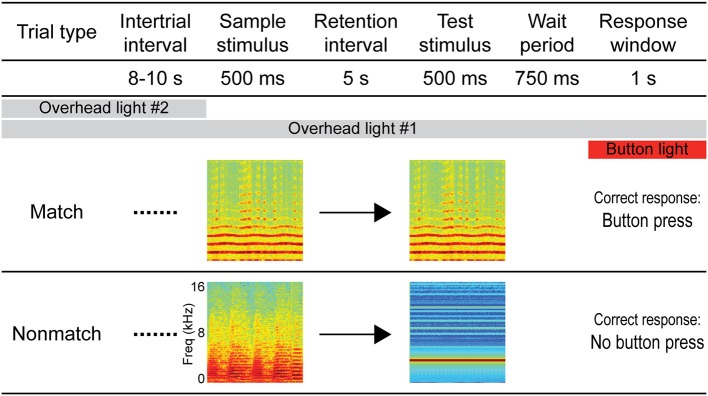
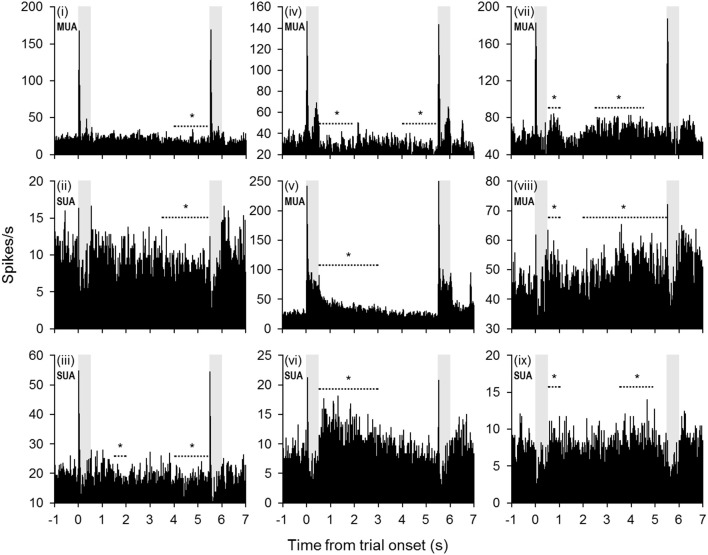
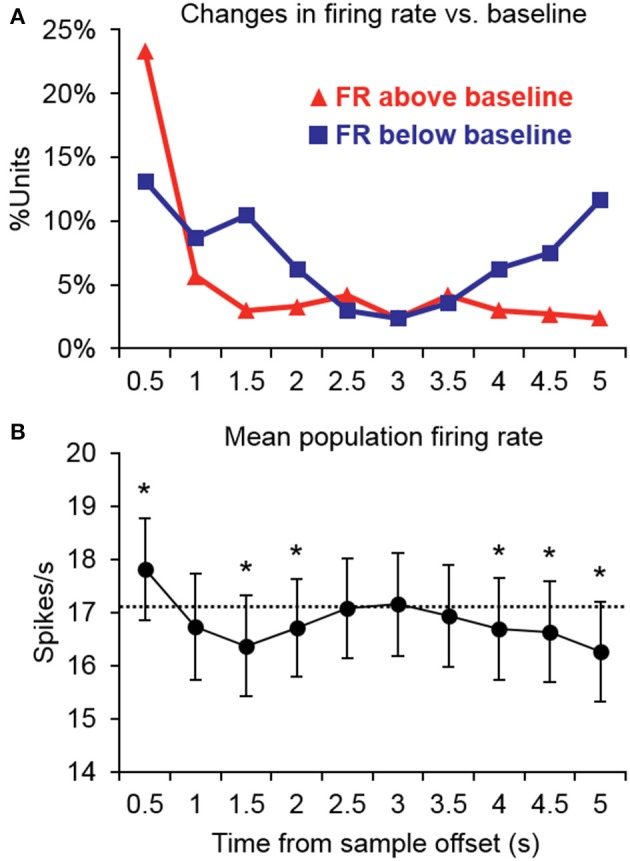
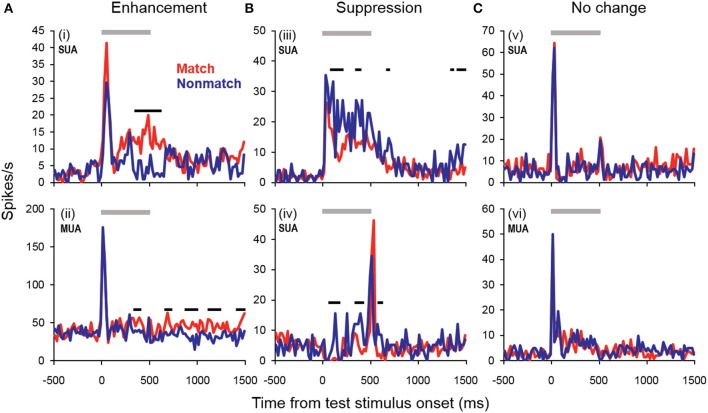
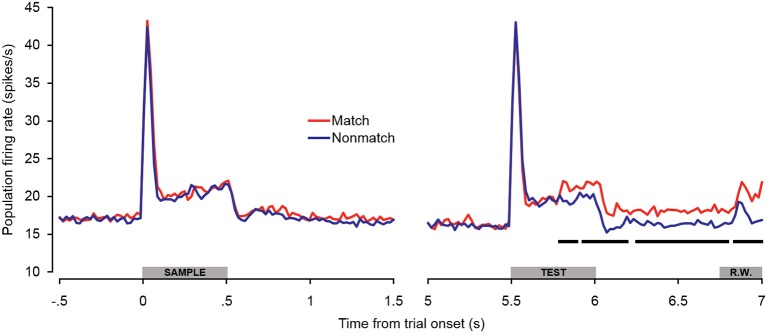
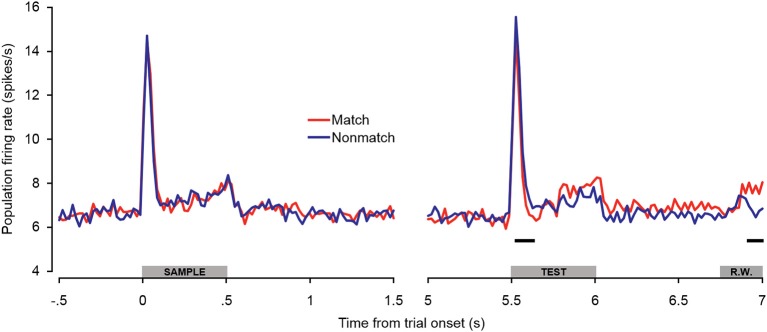
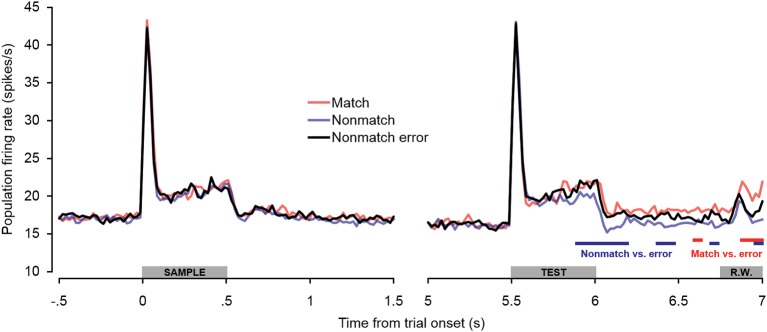
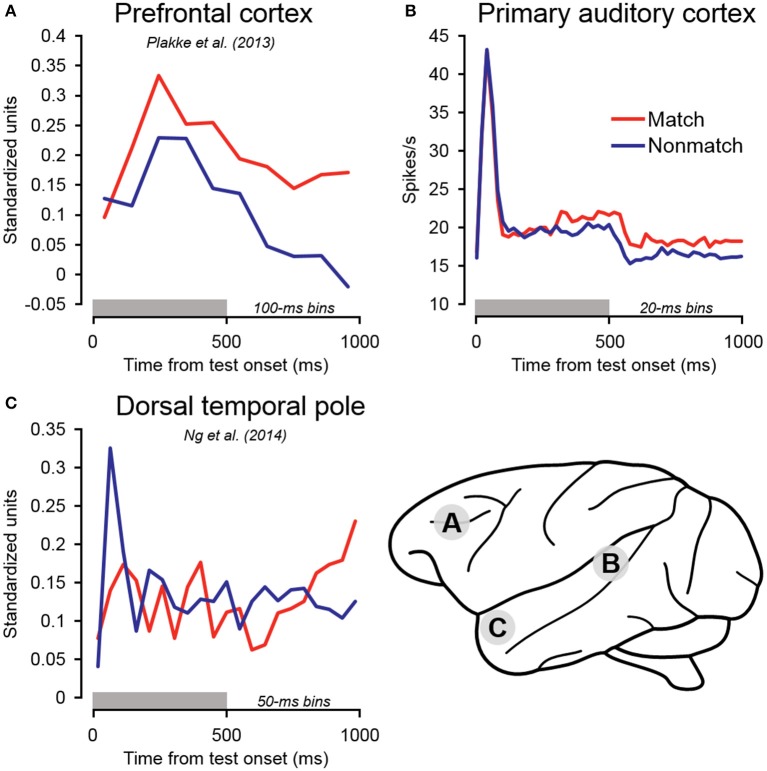
Behaviorally-relevant sounds such as conspecific vocalizations are often available for only a brief amount of time; thus, goal-directed behavior frequently depends on auditory short-term memory (STM). Despite its ecological significance, the neural processes underlying auditory STM remain poorly understood. To investigate the role of the auditory cortex in STM, single- and multi-unit activity was recorded from the primary auditory cortex (A1) of two monkeys performing an auditory STM task using simple and complex sounds. Each trial consisted of a sample and test stimulus separated by a 5-s retention interval. A brief wait period followed the test stimulus, after which subjects pressed a button if the sounds were identical (match trials) or withheld button presses if they were different (non-match trials). A number of units exhibited significant changes in firing rate for portions of the retention interval, although these changes were rarely sustained. Instead, they were most frequently observed during the early and late portions of the retention interval, with inhibition being observed more frequently than excitation. At the population level, responses elicited on match trials were briefly suppressed early in the sound period relative to non-match trials. However, during the latter portion of the sound, firing rates increased significantly for match trials and remained elevated throughout the wait period. Related patterns of activity were observed in prior experiments from our lab in the dorsal temporal pole (dTP) and prefrontal cortex (PFC) of the same animals. The data suggest that early match suppression occurs in both A1 and the dTP, whereas later match enhancement occurs first in the PFC, followed by A1 and later in dTP. Because match enhancement occurs first in the PFC, we speculate that enhancement observed in A1 and dTP may reflect top-down feedback. Overall, our findings suggest that A1 forms part of the larger neural system recruited during auditory STM.
Keywords: A1; Macaca mulatta; primary auditory cortex; recognition memory; rhesus macaque; working memory.
Figures
References
-
- Benjamin A. S., Bjork R. A. (1996). Retrieval fluency as a metacognitive index, in Implicit Memory and Metacognition, ed Reder L. M. (Mahwah, NJ: Earlbaum; ), 309–338
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous