

Effector bottleneck: microbial reprogramming of parasitized host cell transcription by epigenetic remodeling of chromatin structure
- PMID: 25177343
- PMCID: PMC4132484
- DOI: 10.3389/fgene.2014.00274
Effector bottleneck: microbial reprogramming of parasitized host cell transcription by epigenetic remodeling of chromatin structure
Abstract
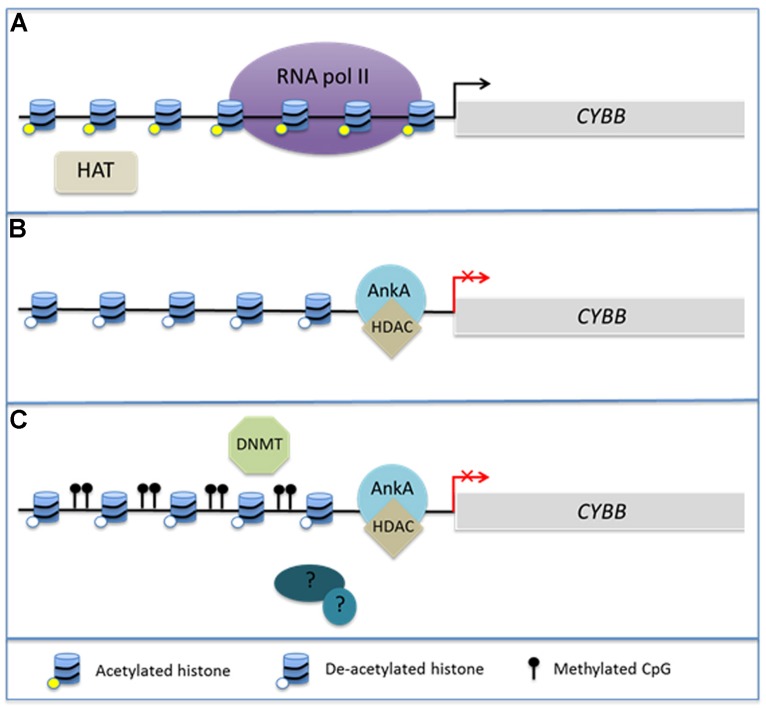
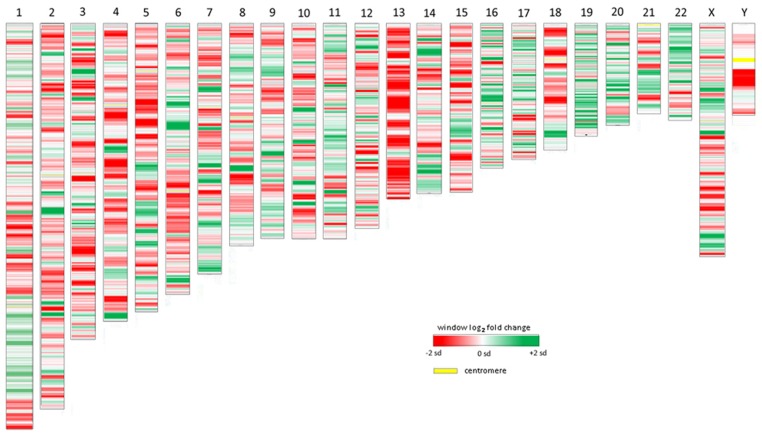
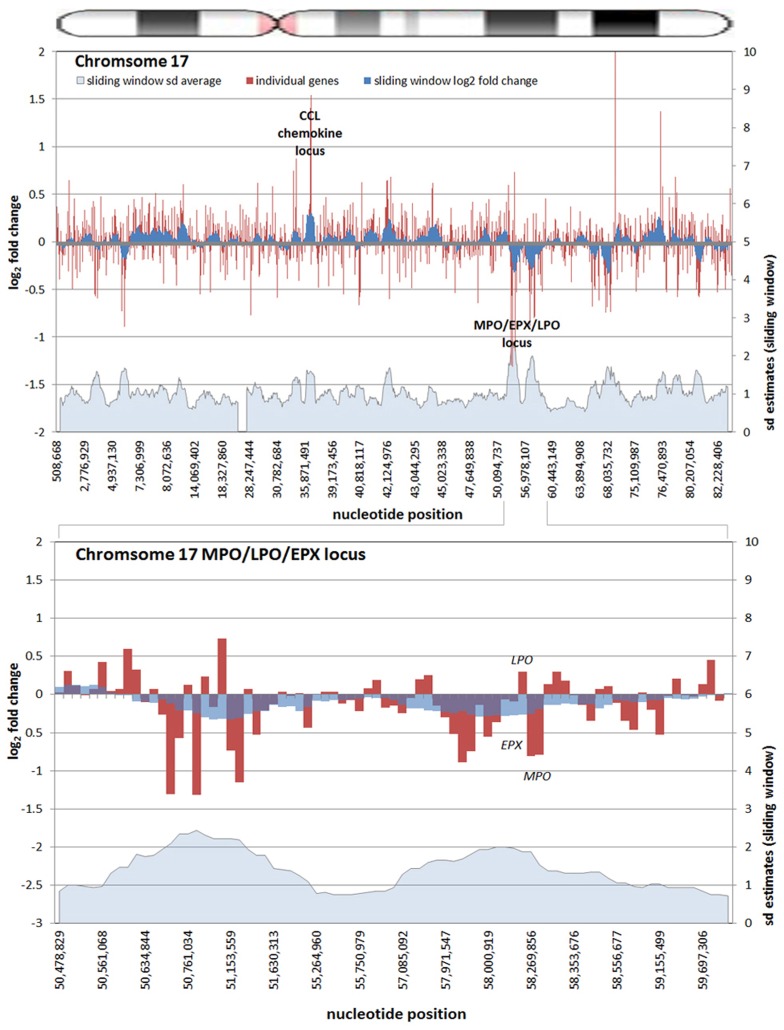
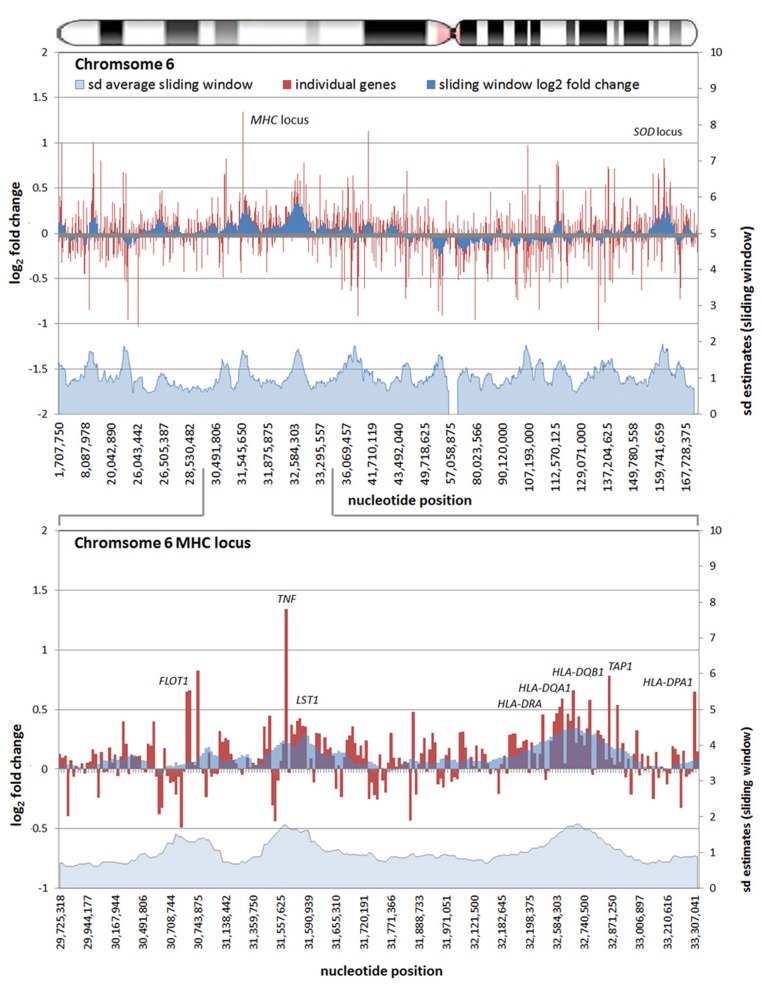
Obligate intracellular pathogenic bacteria evolved to manipulate their host cells with a limited range of proteins constrained by their compact genomes. The harsh environment of a phagocytic defense cell is one that challenges the majority of commensal and pathogenic bacteria; yet, these are the obligatory vertebrate homes for important pathogenic species in the Anaplasmataceae family. Survival requires that the parasite fundamentally alter the native functions of the cell to allow its entry, intracellular replication, and transmission to a hematophagous arthropod. The small genomic repertoires encode several eukaryotic-like proteins, including ankyrin A (AnkA) of Anaplasma phagocytophilum and Ank200 and tandem-repeat containing proteins of Ehrlichia chaffeensis that localize to the host cell nucleus and directly bind DNA. As a model, A. phagocytophilum AnkA appears to directly alter host cell gene expression by recruiting chromatin modifying enzymes such as histone deacetylases and methyltransferases or by acting directly on transcription in cis. While cis binding could feasibly alter limited ranges of genes and cellular functions, the complex and dramatic alterations in transcription observed with infection are difficult to explain on the basis of individually targeted genes. We hypothesize that nucleomodulins can act broadly, even genome-wide, to affect entire chromosomal neighborhoods and topologically associating chromatin domains by recruiting chromatin remodeling complexes or by altering the folding patterns of chromatin that bring distant regulatory regions together to coordinate control of transcriptional reprogramming. This review focuses on the A. phagocytophilum nucleomodulin AnkA, how it impacts host cell transcriptional responses, and current investigations that seek to determine how these multifunctional eukaryotic-like proteins facilitate epigenetic alterations and cellular reprogramming at the chromosomal level.
Keywords: Anaplasma phagocytophilum; DNA methylation; chromatin; epigenetics; histone deacetylase; nucleomodulin.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials