

Cloning and functional analysis of three diacylglycerol acyltransferase genes from peanut (Arachis hypogaea L.)
- PMID: 25181516
- PMCID: PMC4152018
- DOI: 10.1371/journal.pone.0105834
Cloning and functional analysis of three diacylglycerol acyltransferase genes from peanut (Arachis hypogaea L.)
Abstract
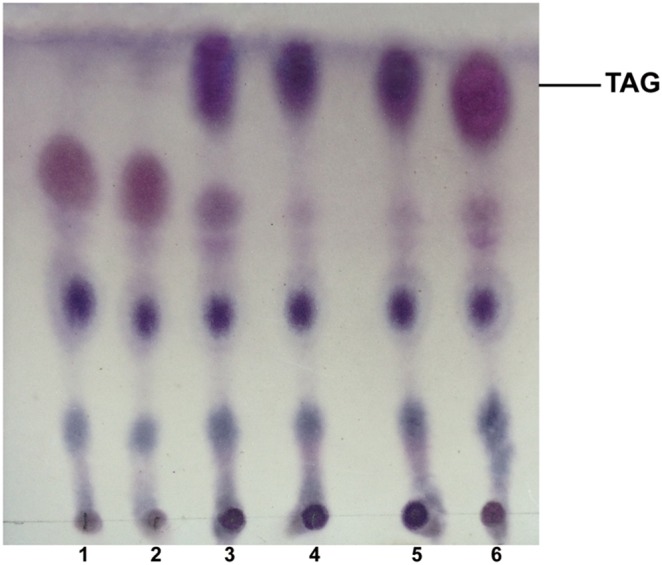
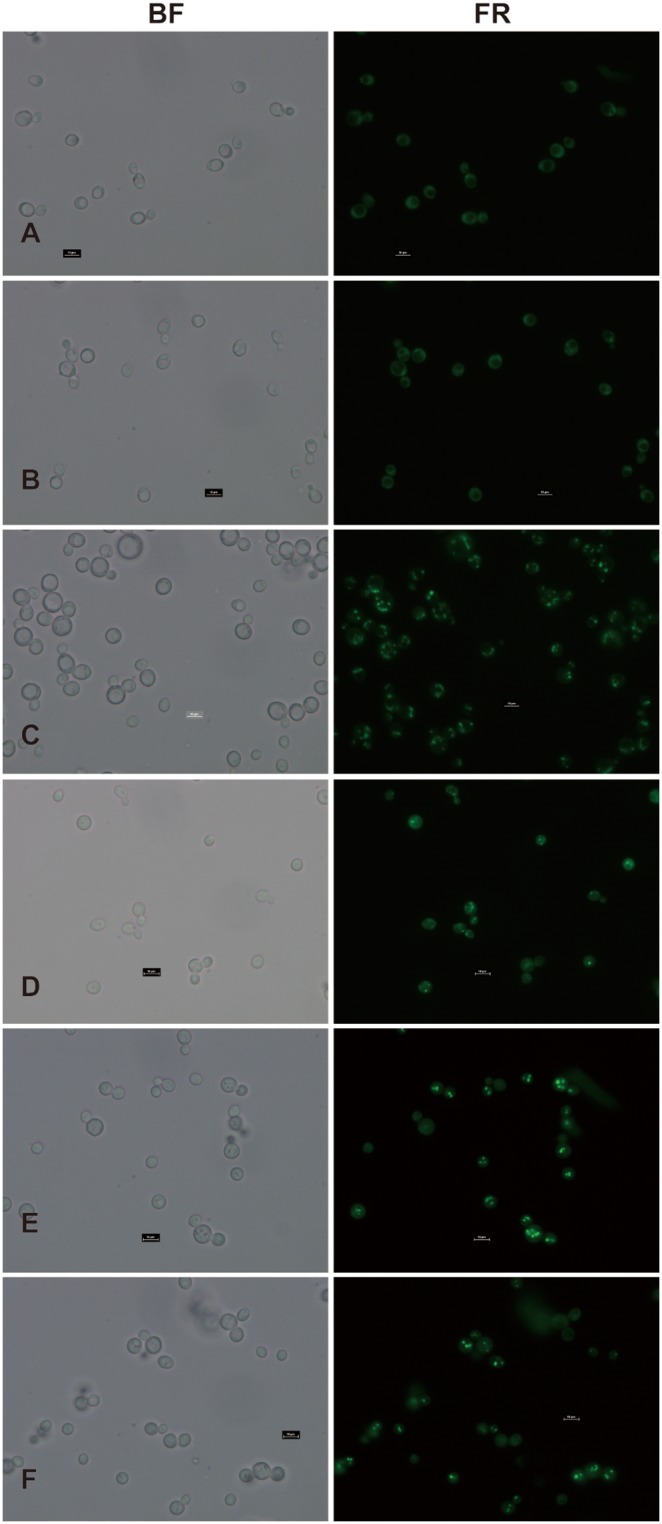
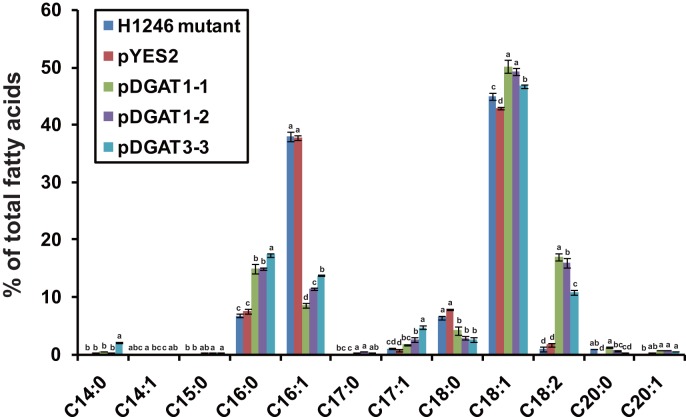
Diacylglycerol acyltransferase (DGAT) catalyzes the final and only committed acylation step in the synthesis of triacylglycerols. In this study, three novel AhDGATs genes were identified and isolated from peanut. Quantitative real-time RT-PCR analysis indicated that the AhDGAT1-2 transcript was more abundant in roots, seeds, and cotyledons, whereas the transcript abundances of AhDGAT1-1 and AhDGAT3-3 were higher in flowers than in the other tissues examined. During seed development, transcript levels of AhDGAT1-1 remained relatively low during the initial developmental stage but increased gradually during later stages, peaking at 50 days after pegging (DAP). Levels of AhDGAT1-2 transcripts were higher at 10 and 60 DAPs and much lower during other stages, whereas AhDGAT3-3 showed higher expression levels at 20 and 50 DAPs. In addition, AhDGAT transcripts were differentially expressed following exposure to abiotic stresses or abscisic acid. The activity of the three AhDGAT genes was confirmed by heterologous expression in a Saccharomyces cerevisiae TAG-deficient quadruple mutant. The recombinant yeasts restored lipid body formation and TAG biosynthesis, and preferentially incorporated unsaturated C18 fatty acids into lipids. The present study provides significant information useful in modifying the oil deposition of peanut through molecular breeding.
Conflict of interest statement
Figures


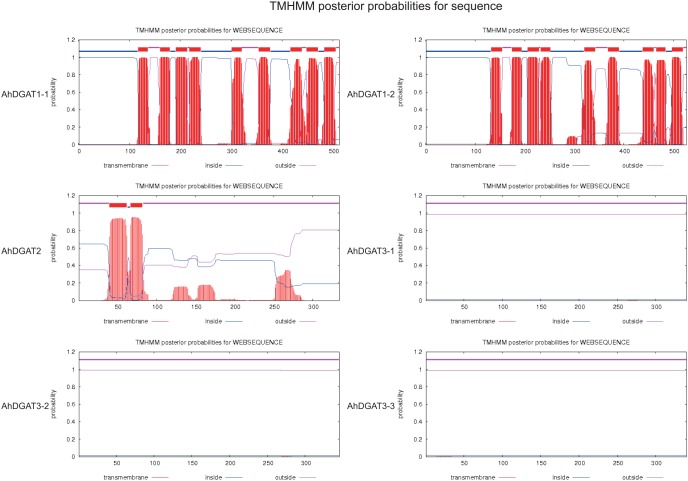
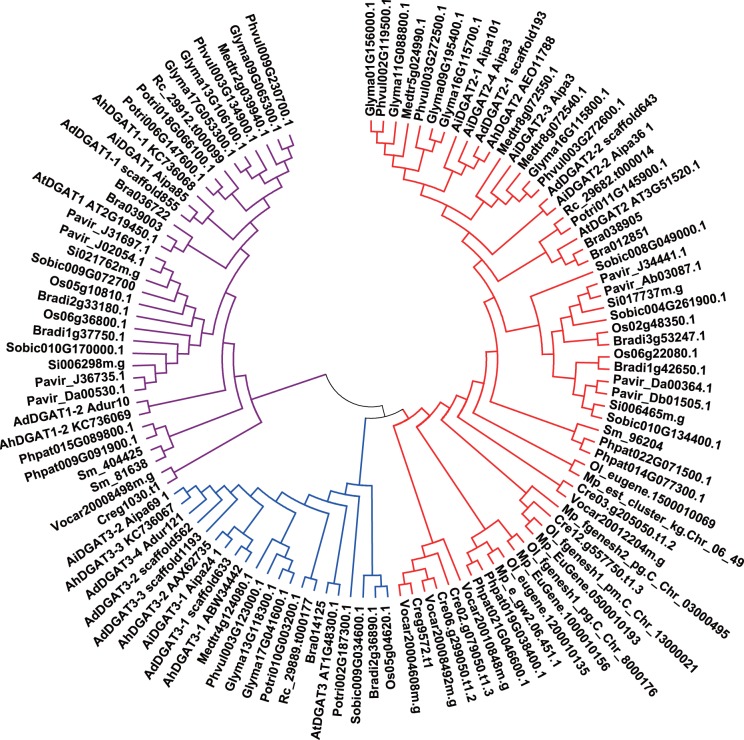
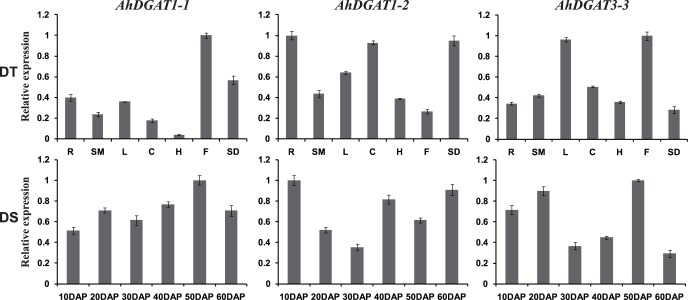
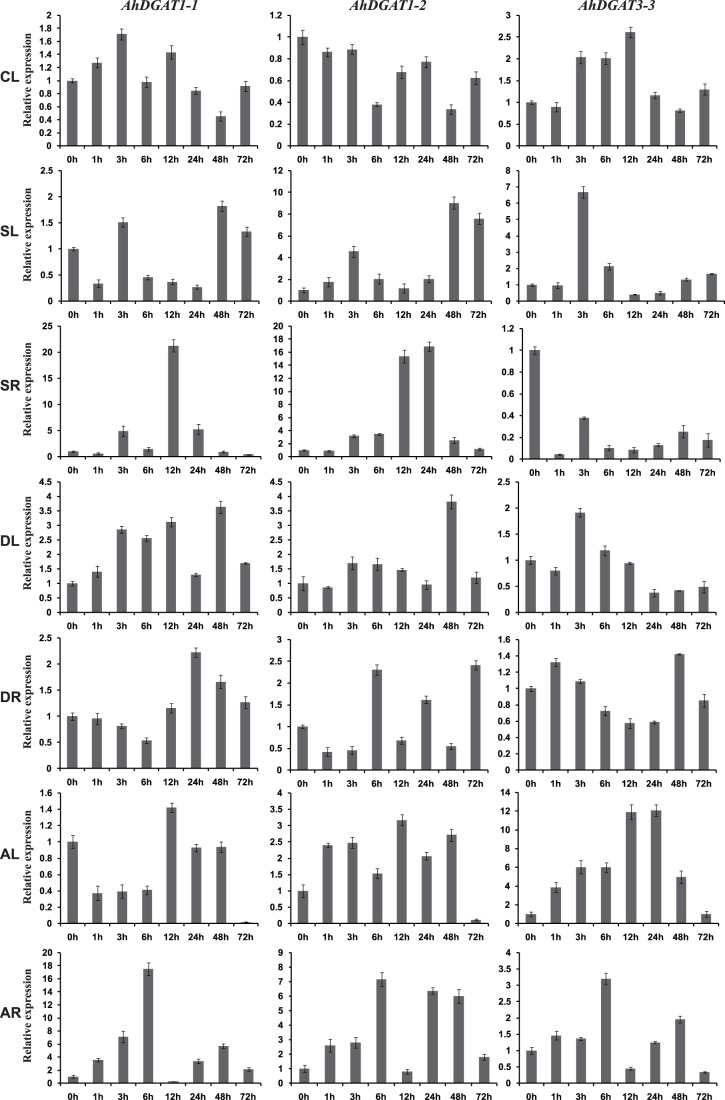
References
-
- Zhang FY, Yang MF, Xu YN (2005) Silencing of DGAT1 in tobacco causes a reduction in seed oil content. Plant Sci 169: 689–694.
-
- Cases S, Novak S, Zheng YW, Myers HM, Lear SR, et al. (1998) ACAT-2, a second mammalian acyl-CoA/cholesterol acyltransferase. Its cloning, expression, and characterization. J Biol Chem 273: 26755–26764. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
